
Streblosoma bairdi ( Malmgren, 1866 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4544.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:E94AAFC5-B0E3-4EEB-ACCF-A5C76439A654 |
|
DOI |
https://doi.org/10.5281/zenodo.5926110 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE6706-4273-FF8B-B1CF-711CEC16F90C |
|
treatment provided by |
Plazi |
|
scientific name |
Streblosoma bairdi ( Malmgren, 1866 ) |
| status |
|
Streblosoma bairdi ( Malmgren, 1866) View in CoL
( Figs 1–2 View FIGURE 1 View FIGURE 2 )
Grymaea bairdi Malmgren, 1866: 388 View in CoL -389, Pl. XIX, Fig. 69A–D.
Streblosoma bairdi View in CoL . Holthe 1986a: 142 –143, Fig. 64a–d. Jirkov 2001: 525, Fig. Streblosoma bairdi View in CoL .
Material examined. Syntypes ( NRM 6491 View Materials ): 2 specs and a mid-body fragment with 8 “thoracic” segments, coll. Sweden, Bohuslän , 58 o 30’N 11 o 10’E, by S. Lovén; both specs incomplete, in poor state of preservation, with notopodia until end of fragments; longest spec. ~ 55 mm long, ~4.5 mm wide, 29 segs, missing most branchial filaments (except for 1–2 at each side), full of oocytes; shortest spec. ~ 27 mm long, 3 mm wide, 23 segs. Slides from last segment of mid-body fragment GoogleMaps .
Additional material examined. NRM 104138 View Materials : coll. Sweden, Bohuslän, Kosterrännan, outside L. Sneholm, by Kristineberg Zoological Station , 0 9 Sep 1924, 100 m, rocks and mud; 2 incomplete specs, in very good state of preservation, one of which with 27 segs, the other with 26, both with notopodia until last segment .
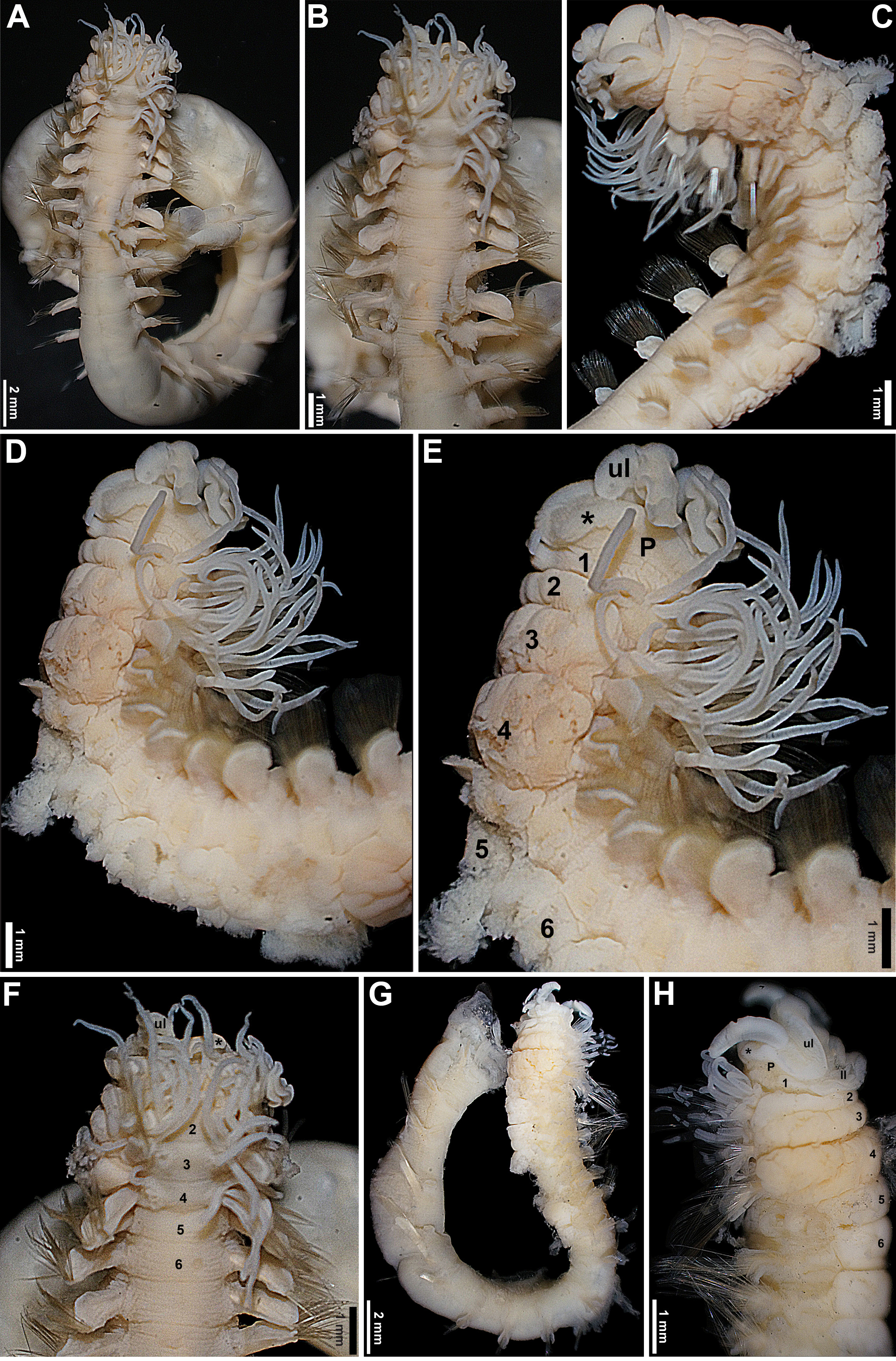
Description. Prostomium at the base of the upper lip; distal part with developed shelf-like process ( Fig. 1 View FIGURE 1 C–H), eyespots on basal part, if originally present, completely faded (including non-types). Peristomium restricted to lips; upper lip short, hood-like, broader than high, convoluted ( Fig. 1 View FIGURE 1 C–F, H); rectangular lower lip, short, distinctly wider than long, with straight lateral and upper margins, rounded at corners, not extending laterally beyond the mouth ( Fig. 1H View FIGURE 1 ). Segment 1 short, more developed ventrally, with lobe marginal to mouth ( Fig. 1 View FIGURE 1 C–E, H); segment 4 longest ( Fig. 1 View FIGURE 1 C–E, G–H). Lateral lobes absent, but segments 2–4 with thickened anterior margins ventrally, as protruding crests ( Fig. 1 View FIGURE 1 C–E, H). Three pairs of branchiae, on segments 2–4, each with ~8–10 independent distally pointed cylindrical filaments, originating from glandular cushion-like structures, with wide medial gap, equivalent to both glandular areas of each pair combined; longest branchial filaments about as long as body width at corresponding segment; on segment 2, branchial filaments arranged in 2 irregular rows, from anterior margin of segment until posterior margin of notopodia, filaments of first row extending laterally until level of notopodia, or very slightly beyond ( Fig. 1 View FIGURE 1 A–H). Ventral surface of anterior segments strongly glandular between parapodia, from segment 2 to ~11, highly corrugated, discrete ventral shields absent ( Fig. 1 View FIGURE 1 C–E, G–H); mid-ventral stripe beginning from ~segment 12. Notopodia starting from segment 2 and extending until end of all fragments, longest specimen with 29 segments; first pair of notopodia shorter than following pairs, except on short specimen; first 2 pairs of notopodia at posterior part of corresponding segments, from segment 4 notopodia at mid-length of segments; notopodia large, roughly rectangular, distally rounded, bilobed, chaetae emerging between lobes ( Fig. 1 View FIGURE 1 A–H). Narrowly-winged notochaetae in both rows, wings only present at tips, those of anterior row much shorter than of those from posterior row, ~1/5 of their length, or less ( Fig. 2 View FIGURE 2 A–C). Neuropodia starting from segment 5, as fleshy ridges, rounded at tips; tori subdistal, nearer to distal ends of neuropodia, but far from tips. Uncini with 3 rows of secondary teeth, teeth progressively shorter upwards, distal row with tiny, almost inconspicuous teeth; dorsal button far from anterior margin, conspicuous prow, longer than button, conspicuous heel, base not as curved as in other species ( Fig. 2 View FIGURE 2 D–F). Nephridial papillae not visible. Pygidium unknown.
Variation. In the shortest syntype, the mid-ventral stripe apparently extends from segment 2, as a continuous groove between segments, among the corrugations of the glandular body wall, and the first pair of notopodia is about the same size as following pairs, instead of shorter, as in the other specimens.
Remarks. The redescription provided by Jirkov (2001) is very brief and mentions very few characters, making the comparison with the Swedish material more difficult. Even though, the morphology of the branchiae as described by Jirkov (2001) raises some questions as if the material examined by the author really belongs to S. bairdi . Jirkov said that the second and the third pairs of branchiae have at least 4 filaments at each side and that these originate directly from the body surface, while the Swedish specimens have almost twice that number of branchial filaments on each side of second and third pairs of branchiae, and those originate from cushion-like, apparently glandular areas. In addition, the drawing of the uncinus provided by Jirkov (2001) shows the dorsal button more away from the anterior margin of the uncinus than in Swedish specimens.
The redescription by Holthe (1986a) was based on complete specimens, and thus the author was able to include characters such as the presence of up to 90 pairs of notopodia and pygidial papillae in his redescription. However, Holthe (1986) did not specify from which locality in Scandinavia came the material he examined, his redescription does not describe in detail characters such as the morphology of the lips, branchiae and notochaetae, and yet there are some discrepancies between his redescription and that provided above. Holthe’s specimens have fewer branchial filaments, at least 7 pairs on segment 2 and 4 pairs on segments 3 and 4, and they have nephridial and genital papillae on segments 3–7, which is rather unusual for a thelepodid, as those papillae, if visible, are always on segments 4–7, among all the animals belonging to this family ( Nogueira et al. 2010, 2013, 2017). In contrast, the syntypes and the specimens from the type locality examined for the redescription provided above have ~8–10 branchial filaments on each pair and nephridial and genital papillae are not visible.
Distribution. Animals from many localities around the world were identified as belonging to S. bairdi , but it is recommended that material from out of the type locality region, especially that from out of the Scandinavian Peninsula, is reviewed to confirm the species identity. Currently, members of this species have been reported from the northern and eastern Atlantic, extending through Cape Verde islands, including the Artcic, and the Mediterranean; Caribbean and southwestern Atlantic, off northeastern to southeastern Brazil and South Georgia; Antarctica; and northern Pacific, along the northern American coast up to the Sea of Okhotsk, and Japan ( Holthe 1986a, b).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Streblosoma bairdi ( Malmgren, 1866 )
| Nogueira, João Miguel De Matos 2019 |
Streblosoma bairdi
| Jirkov, I. A. 2001: 525 |
| Holthe, T. 1986: 142 |
Grymaea bairdi Malmgren, 1866 : 388
| Malmgren, A. J. 1866: 388 |