
Araeosoma bakeri, Anderson, Owen F., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3609.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:046410E8-079F-4FBF-B0EF-361DFCE92E2C |
|
DOI |
https://doi.org/10.5281/zenodo.5622886 |
|
persistent identifier |
https://treatment.plazi.org/id/5B97CA33-B486-43A1-B5D9-827557DC82E6 |
|
taxon LSID |
lsid:zoobank.org:act:5B97CA33-B486-43A1-B5D9-827557DC82E6 |
|
treatment provided by |
Plazi |
|
scientific name |
Araeosoma bakeri |
| status |
sp. nov. |
Araeosoma bakeri View in CoL sp. nov.
Figures 3 View FIGURE 3 B, 4F, 23–25
Holotype —From east of Great Barrier Island, northeast New Zealand: 36° S, 176° E, 182 m (141 mm TD), Te Papa EC1233. Deposited in Te Papa, Wellington. Caught in 1964 by R. B. Pike. Stored dry.
Paratypes —One specimen from Tuatoru Knoll, 37° 28.3' S, 177° 12.9' E, 180– 177 m, NIWA 6564, 144 mm TD, stored in 80% ethanol; six specimens from Tuatoru Knoll, 37° 28.6' S, 177° 12.9' E, 180– 179 m, 134–152 mm TD, NIWA6570, stored in 80% ethanol.
Other material —West Norfolk Ridge: 8 specimens, 32° 39.1', 167° 40.4' E, Te Papa EC3685 (dry); 1 specimen, 32° 41.3 S, 167° 38.1' E, Te Papa EC3684 (dry). Northland: 4 specimens, 35° 19.0' S, 175° 2.0' E, Te Papa EC3691 (dry); 1 specimen, 35° 29.0' S, 175° 2.0' E, Te Papa EC1672 (dry); 2 specimens (87, 143 mm TD), 35° 39.7' S, 175° 33.2' E, 348, NIWA29448; 2 specimens (134, 120 mm TD), 36° 13.0' S, 176° 12.7' E, 343– 336 m, NIWA29523; 1 specimen, 36° 13.9' S, 176° 11.6' E, Te Papa EC7503 (dry); 4 specimens (106, 112, 123, 130 mm TD), 36° 14.0' S, 176° 12.1' E, 340– 340 m, NIWA29447. Bay of Plenty: 2 specimens, 37° 21.4' S, 177° 5.9' E, Te Papa EC3682 (dry); 1 specimen, 37° 16.2' S, 176° 50.6' E, Te Papa EC3681 (dry); 1 specimen (130 mm TD), 37° 15.1' S, 176° 50.1' E, 195 m, NIWA23716; 1 specimen (74 mm TD), 37° 16.6' S, 176° 50.1' E, 330 m, NIWA29437 (dry); 3 specimens, 37° 17.4' S, 176° 53.6' E, Te Papa EC9362; 3 specimens, 37° 17.4' S, 176° 53.6' E, Te Papa EC4115; 3 specimens (frozen), 37° 19.8' S, 177° 4.4' E, 358–380 m, NIWA6575; 7 specimens (frozen), 37° 19.9' S, 177° 4.5' E, 375–414 m, NIWA6578; 1 specimen (frozen), 37° 20.4' S, 177° 6.7' E, 335– 275 m, NIWA6576; 2 specimens, 37° 20.5' S, 177° 5.4' E, Te Papa EC9467; 2 specimens (frozen), 37° 21.0' S, 177° 6.0' E, 434– 272 m, NIWA6585; 18 specimens (frozen), 37° 21.3' S, 177° 6.0' E, 260–280 m, NIWA6577; 31 specimens (frozen), 37° 21.3' S, 177° 6.1' E, 259–294 m, NIWA6579; 2 specimens (frozen), 37° 27.5' S, 176° 54.1' E, 312– 278 m, NIWA6580; 3 specimens (frozen), 37° 27.6' S, 176° 55.6' E, 320– 302 m, NIWA6583; 3 specimens (frozen), 37° 27.7' S, 176° 54.8' E, 294– 247 m, NIWA6584; 1 specimen (discarded), 37° 28.0' S, 176° 54.9' E, 225– 210 m, NIWA6582; 1 specimen, 37° 28.1' S, 176° 55.8' E, 315– 283 m, NIWA6581; 6 specimens (104, 129, 132, 130, 132, 139 mm TD), 37° 28.2' S, 177° 13.2' E, 200– 175 m, NIWA6563; 1 specimen (135 mm TD), 37° 28.2' S, 177° 7.0' E, 220–320 m, NIWA29424; 20 specimens, 37° 28.2' S, 177° 13.4' E, 207–216 m, NIWA6568 (discarded) & NIWA6569; 11 specimens (frozen), 37° 28.3' S, 177° 13.5' E, 218– 200 m (NIWA6565, NIWA6566, NIWA6567, & NIWA6588); 1 specimen, 37° 28.3' S, 177° 12.9' E, Te Papa EC9522; 2 specimens (35, 127 mm TD), 37° 28.5' S, 176° 54.5' E, 286– 238 m, NIWA6606; 1 specimen (frozen), 37° 32.0' S, 176° 58.8' E, 219– 176 m, NIWA6572; 1 specimen (123 mm TD), 37° 33.4' S, 176° 58.9' E, 266– 197 m, NIWA6571; 1 specimen (128 mm TD), 37° 33.5' S, 176° 58.9' E, 210–314 m, NIWA29457; 8 specimens (frozen), 37° 33.6' S, 176° 59.1' E, 313–314 m, NIWA6573 & NIWA6574; 2 specimens (119, 129 mm TD), 37° 33.8' S, 176° 58.7' E, 370– 176 m, NIWA29418. East Coast North Island: 1 specimen (118 mm TD), 39° 45.8' S, 178° 14.1' E, 288 m, NIWA29428. Unknown locality: 6 specimens (130, 127, 132, 145, 125, 124 mm TD), NIWA23709 (dry). Unless indicated, stored in 80% ethanol.
Note: Baker’s other specimen (No. 2), collected from the Bay of Plenty in 163–182 m, NMNZ 1232, was neither A. bakeri nor A. coriaceum but in fact Hapalosoma amynina sp. nov.
Etymology —Named bakeri , after Alan Baker, who provided the initial description of the species, used as a noun in the genitive case.
Diagnosis —Adults large, sub-pentagonal in outline, up to 152 mm TD; test and spines blackish violet; tridentate pedicellariae of three types, a large involute type with strongly enclosed valves and two smaller rostrate types; dactylous pedicellariae with 4 valves; short triphyllous pedicellariae; primary tubercles with large, overlapping areoles, those in the oral adradial interambulacral series occurring on every plate; oral primary spines with extremely short hoofs.
Description —The holotype and a specimen of Hapalosoma amynina were initially examined by Baker (1972) and mistakenly identified as A. coriaceum . Although there are a number of similarities between A. coriaceum and A. bakeri —mainly in the colour, the shortness of the hoofs, and in some aspects of test morphology—there are distinct differences between the species in the pedicellariae, tuberculation patterns, overall size, and depth range. Along with his recording of this species in northern New Zealand, Baker (1972) provided a valuable description of the holotype, with excellent figures of the pedicellariae, but his drawing of the oral test plates was unfortunately based on the specimen of H. amynina . This figure (his figure 1) shows the more regular inner series of primary tubercles typical of H. amynina (in comparison with Baker’s plate 1 and Figure 32 View FIGURE 32 in this paper). It is now possible to provide some further details of A. bakeri and make a more complete description, having had the opportunity to examine a large number of additional specimens.
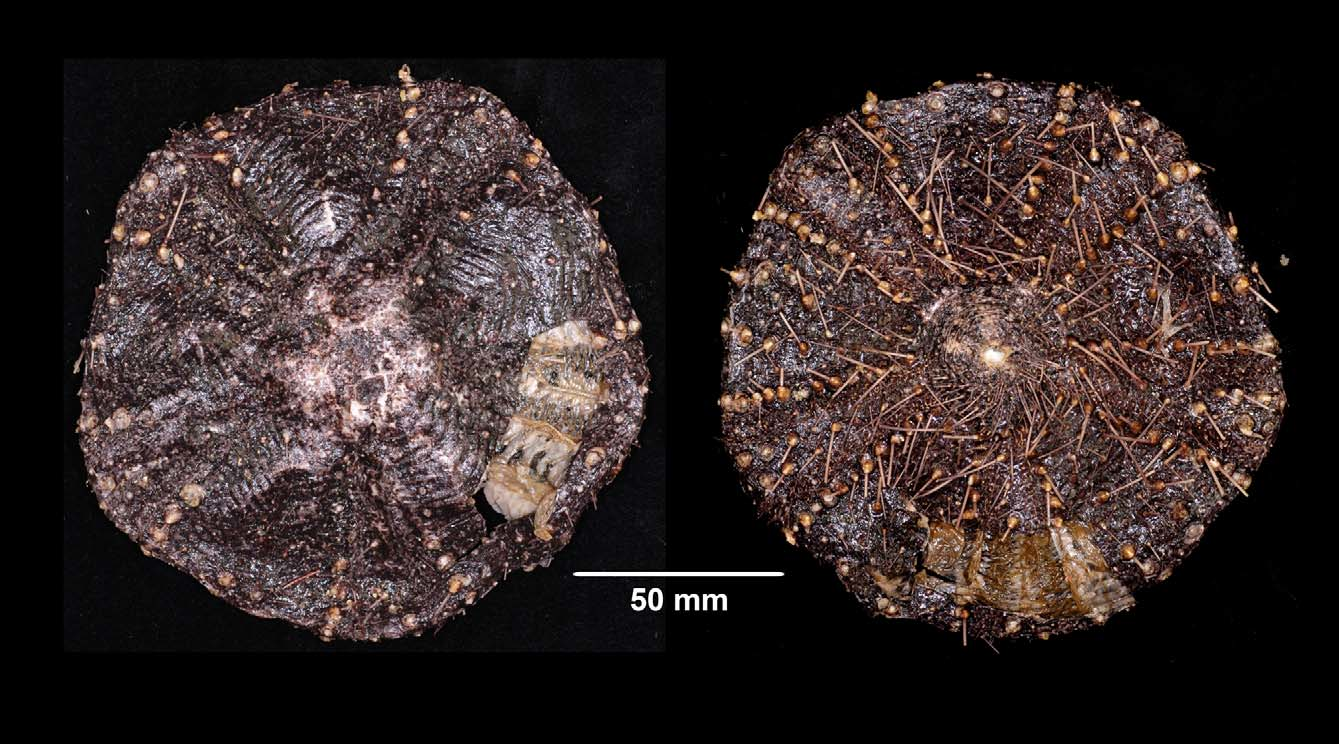
All individuals examined have to some degree a distinct pentagonal outline, rather than circular ( Figure 23 View FIGURE 23 ). The purplish-black colour of the test and spines seen by Baker is also consistent among the additional specimens examined. The size of Bakers specimen (141 mm TD) was typical; the specimens now measured ranging from 87 to 152 mm TD. A distinctive feature of the species is in the areoles of the primary tubercles. These are particularly large and frequently extend beyond the edge of the plate, with those of the main oral interambulacral series overlapping each other considerably.
Apical system typical of the genus and made up of many small plates, as described by Baker. In the specimens examined there is quite some variability in the shape of the overall outline of the apical system, the genital plates in a few being much extended down the midline of the interambulacral columns, with the result that the upper plates of these columns must angle and bend around them, causing the “herring-bone” pattern noted by Baker and common in Araeosoma . In other specimens genital plates more compact and interambulacral plates much less angled. Genital pore opens into a membranous space within the disaggregated genital plates, rather than beyond them as is often reported for this genus, with one or two small plates clearly discernible beyond the pore, even in wet specimens. Madreporite scarcely raised—varying from kidney-shaped to heartshaped to triangular. Ocular plates with 2–3 small tubercles, the pore at the outer edge. Numerous small plates cover remainder of apical system, each bearing a few small tubercles. Anus central, surrounded by a rough ring of tubercles.
Number of plates per column relatively consistent between specimens. Orally there are 14–16 interambulacral and 20–23 ambulacral plates; aborally 20–24 interambulacral and 30–35 ambulacral plates. As several dry specimens were available it was possible to determine the plate boundaries relatively easily. Membranous tissue forms wide gaps between successive plates in the interambulacra, particularly aborally. These gaps variable between specimens on the oral surface, from being almost non-existent in some to being as much as about half the mid-plate width in others. Plates in the ambulacra generally closely abutting.
To Baker’s (1972) description of the oral tuberculation can be added that in the interambulacra there are usually one or two primary tubercles in the outer plates, between the median series and the midline. The variability noted by Baker in the ambulacral tuberculation is also seen in the additional specimens examined, the primary tubercles variously occurring on adjacent or every second or third plates.
The continuation of the oral ambulacral and adradial interambulacral series of tubercles onto the aboral side (for 1–2 plates and 4–5 plates respectively), as noted by Baker, is quite a consistent feature among the specimens examined. In particular, the large tubercles of the interambulacral series are a distinctive feature of the aboral side as they reach sometimes to over a third of way up the column towards the apical system, occasionally missing a plate. There is also an indistinct but consistent series of tubercles in the aboral interambulacra, occurring on most plates, in about mid-plate position near the ambitus but becoming closer to the adradial edge adapically. These tubercles variable in size, the areoles of some encompass the entire height of the plate.
Pore-pairs of the primary ambulacral plates are located at their adapical, adambulacral corner but slightly further perradial near the ambitus. Pore-pairs of the accessory plates lie in close pairs (in the manner described above for A. leppienae ), near the centre of the main plate, slightly separated from its pore-pair.
Peristome about 33 mm in diameter with two overlapping columns of 8–9 slightly angled plates in each ambulacral zone, generally without any additional small plates distally. Each plates bears a single tube-foot and a row of several tubercles spanning the width of the plate. Buccal notches shallow; gills moderate size, lobate.
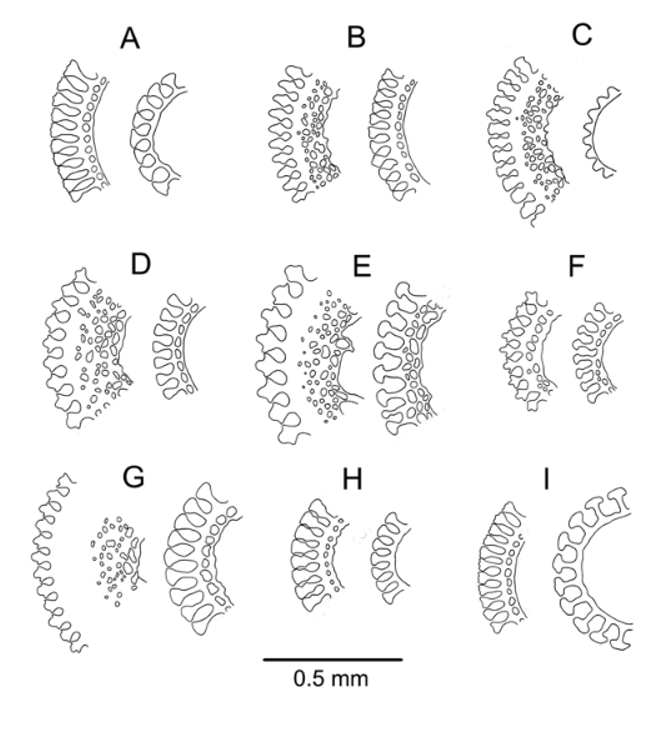
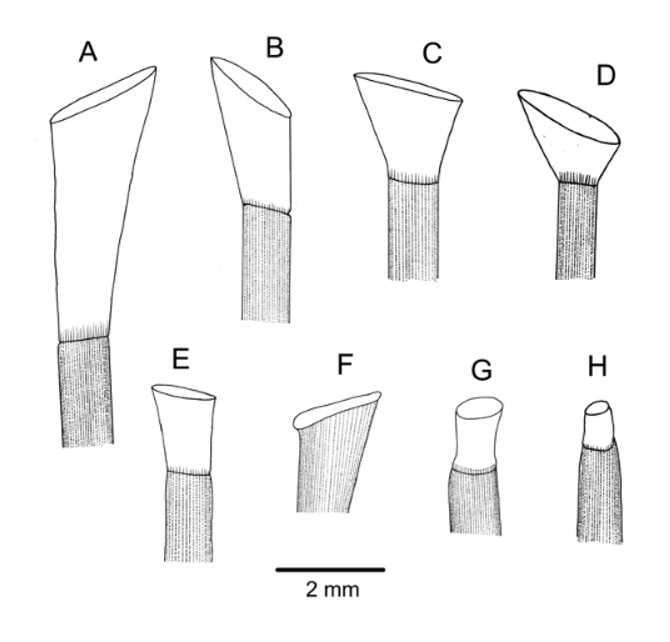
Spines smooth, cylindrical, hollow, fragile, and readily lost on capture; milled ring not prominent, slightly oblique. The largest spines about 25 mm long and 1.0 mm in diameter with about 40 fine, longitudinal striations. These striations simple club-shaped wedges in cross-section in the aboral spines examined, but wedges with distal extensions in the oral spines. Oral spine wall moderately thick, with a layer of meshwork; aboral spine wall thin, enclosing a single row of perforations ( Figure 3 View FIGURE 3 B). Few oral spines with hoofs retained in any specimen; these spines more or less curved and expanded distally, tipped with a very small hoof ( Figure 4 View FIGURE 4 F).
Sphaeridia elongate, hemi-spherical distally (about 0.4 mm long by 0.3 mm wide), located adjacent to the inner tube foot in each ambulacral plate, slightly perradial and adoral to it. None found on the several closest plates to the apical system in the holotype.
The tridentate pedicellariae were well described and figured by Baker and, although not stated, clearly came from his specimen No. 1. Some additional images are provided here of the large involute and smaller tridentate forms originally figured ( Figure 24 View FIGURE 24 ) and of triphyllous and dactylous forms not previously shown, from pedicellariae off paratype NIWA6564. Triphyllous and smaller tridentate pedicellariae were abundant over much of the test on each of the several specimens examined but the large and distinctive involute forms were quite rare, although more numerous on the smaller specimens. The dactylous pedicellariae were rarer still, only a few found on NIWA6564—towards the ambitus on the aboral side. These had 4 valves, about 1.3 mm long. Triphyllous pedicellariae with a head length of about 0.4–0.5 mm, varying in the extent to which the basal part of the valve is completely enclosed. Large involute tridentate pedicellariae with a head length of about 2 mm; small tridentate pedicellariae with a head length of about 0.9–1.1 mm.
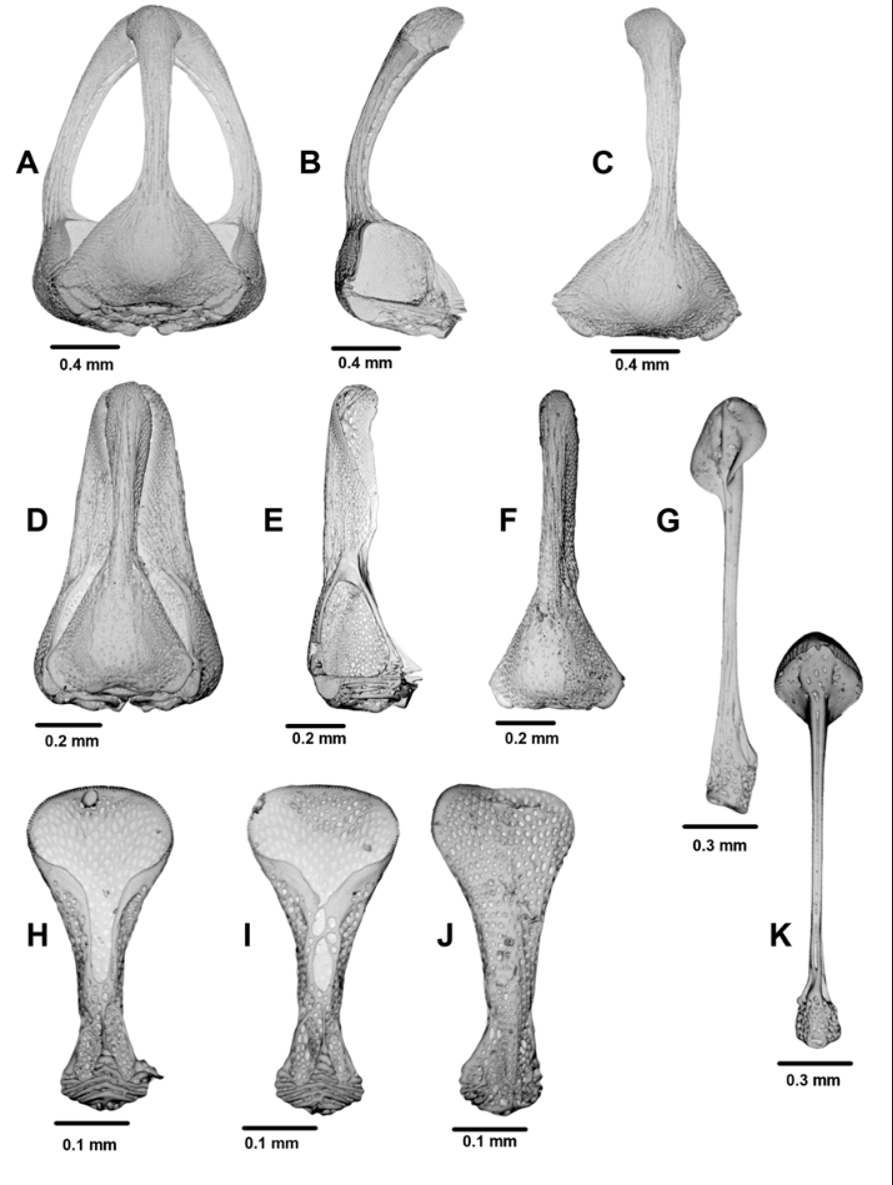
The Aristotle’s lantern has some features which make it stand apart somewhat from the other Araeosoma species covered in this review: the demipyramids are strongly curved and relatively wide for their size; the foramen magnum is very deep, equal to nearly half the length of the lantern; and the epiphysis and brace (rotule) are both relatively long and narrow.
Size range —The median test diameter of the 33 specimens measured was 125 mm, and the largest specimen was 152 mm TD.
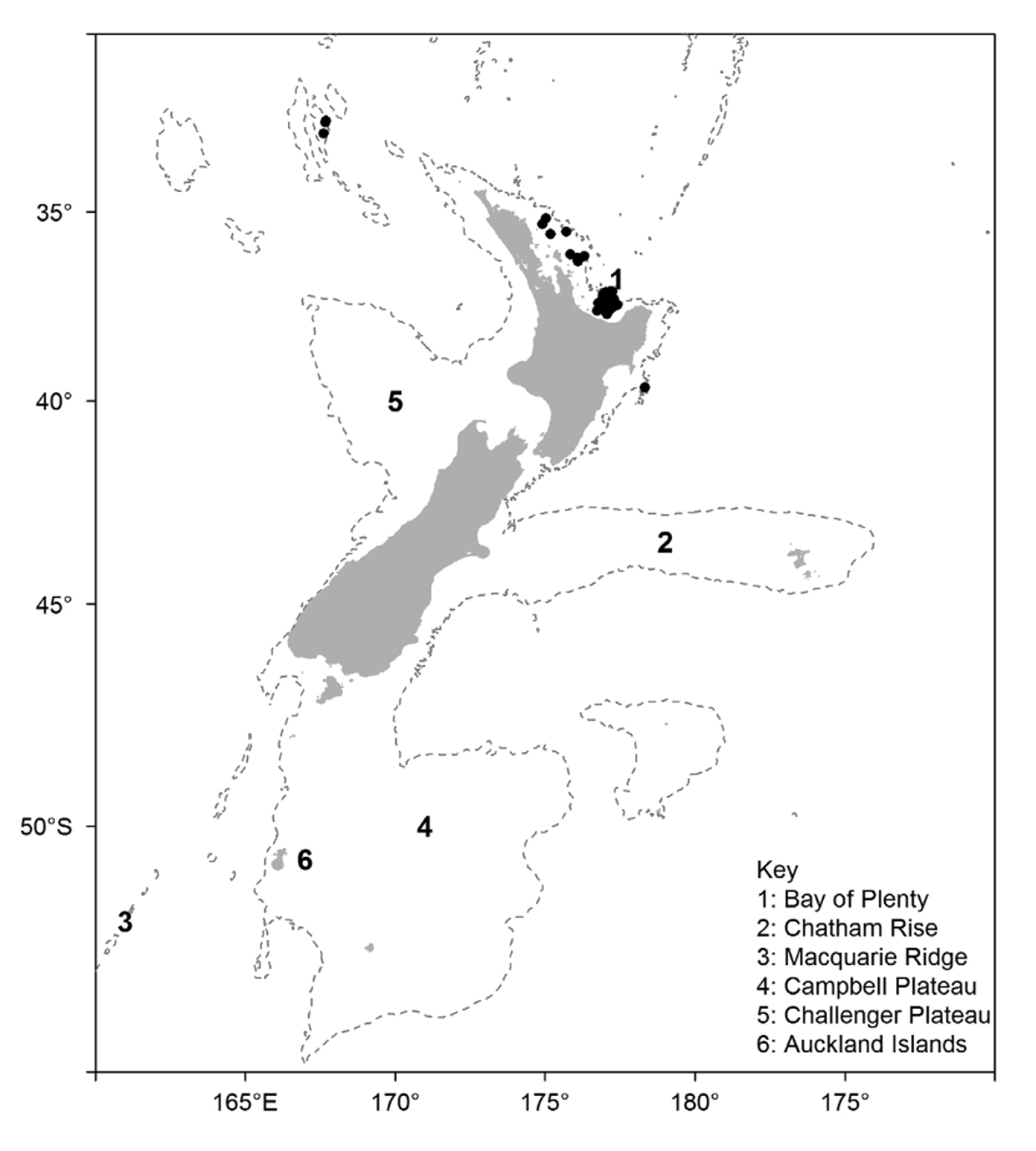
Occurrence —There are 42 records of A. bakeri , representing 176 individuals, all from northern New Zealand ( Figure 25 View FIGURE 25 ). Most records are from the Bay of Plenty and off the north-eastern coast of the North Island, the exceptions being two records from the West Norfolk Ridge and one from off Hawke Bay on the North Island east coast. The species is strongly associated with rocky substrates, with more than half of the records from seamounts sampled on a single RV Tangaroa survey in the Bay of Plenty in 2004. They also occur in relatively high densities, as the average catch for these records was 4 and as many as 31 have been taken in a single trawl. This is unusual, as no more than 6 specimens of any other New Zealand echinothuriinid have been taken at once.
The conservative depth range for the species is 180–375 m, with a potential range of 175– 434 m.
Remarks —With the benefit of having been able to examine a large amount of material it is now clear that this species is distinct from A. coriaceum . Although in his original description Agassiz (1881) provided no figures of the pedicellariae of A. coriaceum , some were later published by Mortensen (1903; 1935; 1940). These show that although the triphyllous pedicellariae are fairly typical for the genus the tridentate pedicellariae (the involute form especially) differ from those of A. bakeri . The dactylous pedicellariae are in both species of the same general form typical of Araeosoma —with four valves (or just as often five in A. coriaceum )—but are far more elongate in A. coriaceum than in A. bakeri .
The tuberculation of the test shows some similarities between the two species, particularly on the oral side (compare Baker (1972) Plate 1 with Mortensen (1935) Plate XXXIII), but the primary tubercles on the aboral side are a continuation of the median oral series in A. coriaceum whereas in A. bakeri they continue the adradial series. In terms of colour, the dark grey, greenish-black, or blackish-brown reported for A. coriaceum differ slightly from the uniform purplish-black of A. bakeri . Orally, although both species have a very short hoof to the primary spines (probably shorter in A. bakeri ) these spines are not widened in A. coriaceum as they are in A. bakeri .
One of Agassiz’ original specimens of A. coriaceum was 220 mm TD (Agassiz 1881), and Agassiz and Clark (1909) also emphasise the large size of this species. Mortensen (1935) describes 3 specimens from different localities, all 190 mm TD. The largest specimen of A. bakeri found is considerably smaller, 152 mm TD.
A similar form of the distinctive involute tridentate pedicellariae of A. bakeri is also found in several other species: A. “ leptaleum ”, A. violaceum , A. tesselatum , A. fenestratum , Calveriosoma gracile (Mortensen 1935) , and A. migratum , and short hoofs are of course a distinctive feature of A. parviungulatum , so it is perhaps among these species (as well as A. coriaceum ) that its closest relatives are to be found. Of these the most similar appears to be A. tesselatum , particularly in the arrangement of large tubercles on the oral surface, but it otherwise differs in the pattern of aboral tuberculation.
It also seems quite certain now that the specimen of A. coriaceum from the Challenger station 169 off NE New Zealand, about which there has been much confusion (see Baker 1972, Mortensen 1935, Clark 1925), was incorrectly labelled. Apart from the morphological differences noted here between that species and A. bakeri , this specimen of A. coriaceum is about 40 mm larger in diameter than any A. bakeri examined and was found in water more than three times the maximum depth recorded for A. bakeri —and deeper than any echinothuriinid record from the New Zealand region. Other confirmed records of A. coriaceum were from about 200 m deeper than A. bakeri has been found. No specimens matching the description of A. coriaceum have been found in the New Zealand region despite considerable sampling effort.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.