
Araeosoma migratum, Anderson, Owen F., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3609.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:046410E8-079F-4FBF-B0EF-361DFCE92E2C |
|
DOI |
https://doi.org/10.5281/zenodo.5622876 |
|
persistent identifier |
https://treatment.plazi.org/id/EBFC7F3F-EE6A-43C5-B523-324407462058 |
|
taxon LSID |
lsid:zoobank.org:act:EBFC7F3F-EE6A-43C5-B523-324407462058 |
|
treatment provided by |
Plazi |
|
scientific name |
Araeosoma migratum |
| status |
sp. nov. |
Araeosoma migratum View in CoL sp. nov.
Figures 3 View FIGURE 3 A, 4D, 7–11
Holotype —From Mt Boyes seamount on the Challenger Plateau, west of New Zealand: 37° 29.0´S, 167° 41.1´E, 581–904 m (140 mm TD), NIWA45089. Deposited in the NIC, Wellington. Collected on 10 September 1998 by Warrick Lyon and Andrew Sealey, government fisheries observers onboard the trawler FV Arrow while bottom trawling for orange roughy, black cardinalfish ( Epigonus telescopus ), and alfonsino ( Beryx splendens ). Stored in 80% ethanol.
Paratypes —Three specimens, all from the southeast Chatham Rise: 1 specimen (125 mm TD), 44° 1.7´S, 174° 35.5´W, 796–882 m, NIWA54300; 1 specimen (127 mm TD), 44° 29.4´S, 175° 5.8´W, 604–831 m, NIWA45095; 1 specimen (128 mm TD), 44° 36.7´S, 175° 16.2´W, 827–1170 m, NIWA45103. All stored in 80% ethanol.
Other material —Northeast Chatham Rise: 1 specimen, 42° 34.3´S, 173° 24.4´W, Te Papa EC9369. Campbell Plateau: 1 specimen, 46° 41.0´S, 165° 43.0´E, Te Papa EC6606; 1 specimen (178 mm TD), 49° 10.9´S, 166° 34.9´E, 631– 610 m, NIWA46090; West Coast South Island: 1 specimen (24 mm TD), 42° 43.0´S, 168° 42.0´E, 1256 m, NIWA34851. East Coast North Island: 1 specimen, 39° 25.8´S, 178° 18.2´E, Te Papa EC9368; 2 specimens (155, 161 mm TD), 40° 2.3´S, 178° 8.6´E, 730–747 m, NIWA45994; 1 specimen (127 mm TD), 39° 32.7´S, 178° 19.8´E, 790–815 m, NIWA45950. Bay of Plenty: 1 specimen (120 mm TD), 37° 2.8´S, 176° 29.8´E, 949 m, NIWA45129; 1 specimen (160 mm TD), 37° 5.1´S, 176° 41.1´E, 985 m, NIWA29524. Challenger Plateau: 1 specimen (115 mm TD), 37° 19.7´S, 168° 2.1´E, 890–937 m, NIWA66070; 1 specimen (157 mm TD), 40° 3.4´S, 167° 57.6´E, 834–1000 m, NIWA34843; 1 specimen (135 mm TD), 37° 30.0´S, 167° 41.0´E, 943 m, NIWA45128. Lord Howe Rise: 1 specimen (127 mm TD), 34° 9.0´S, 162° 51.1´E, 793 m, NIWA29415. West Norfolk Ridge: 1 specimen, 34° 7.0´S, 167° 37.0´E, 870 m, MV (unregistered); 1 specimen, 33° 20.5´S, 170° 14.0´E, 614–675 m, MV F172623; 1 specimen (166 mm TD), 34° 17.1´S, 168° 21.5´E, 785–800 m, MV F172625. Norfolk Ridge: 1 specimen, 26° 25.9´S, 167° 10.9´E, 750–774 m, NIWATAN0308/43. Northland: 1 specimen, 34° 57.8´S, 175° 12.6´E, 614– 602 m, NIWA3478. Tasmania: 1 specimen, 44° 12.6´S, 147° 2.6´E, 1078– 593 m, AM J24542 View Materials ; 20 specimens, 44° 9.1´S, 147° 7.7´E, 800–950 m, AM J24562 View Materials . Unknown locality: 1 specimen (130 mm TD), NIWA23711. All stored in 80% ethanol except for NIWA23711 and Te Papa EC6606 which are stored dry.
Etymology —Named migratum , a past participle used as an adjective, because of the tendency for the distal component of the genital plates to migrate down the interambulacral midline.
Diagnosis —Adults moderately large, up to 166 mm TD; colour of test and appendages brown in various shades; distinct membranous spaces between test plates, particularly orally and in the interambulacra; primary tubercles on every plate in the oral adradial interambulacral series; spines very delicate; hoofs white, short and flaring; large, involute tridentate pedicellariae with a long, narrow, curved blade—only slightly widened at the tip.
Description —Test of holotype ( Figure 7 View FIGURE 7 ) 140 mm TD, circular, flattened, ambitus rounded. Test flexible, moderately robust; the spines fragile, most primary spines missing or broken. Colour of ethanol-preserved test brownish, lighter skin between the plates, darker on the tissue surrounding the tubercles, the spines pale, similar colour to the test. Hoofs short but strongly flared. Photographs taken of a fresh specimen (NIWA45994) showed a similar colouration, except that the tissue surrounding the tubercles was somewhat lighter and there was a reddish tinge to the peristome and pore zones of the ambulacra. Primary tubercles perforate and non-crenulate. Ratio of interambulacrum to ambulacrum width at the ambitus, 3:2.
There are 40 plates in the interambulacral columns cleaned for examination, about 18 on the oral side and 22 on the aboral side. The cleaned ambulacral columns have 61 plates, about 29 on the oral side and 32 on the aboral side. Although difficult to determine exactly without damaging the specimens, plate numbers in the paratypes appeared similar, with about 41–44 plates in the interambulacra and 60–65 plates in the ambulacra in the four specimens examined. Exact plate boundaries partially concealed by thick skin, but wide membranous spaces evident in the median area between successive plates in a column, more apparent aborally and in the interambulacra. Plates of interambulacra and ambulacra oblong, typical for the genus, widened laterally where necessary to accommodate large primary tubercles, the exact boundaries obscured by thick skin.
Oral test plating ( Figure 8 View FIGURE 8 ): A regular series of adradial primary tubercles on every plate in the interambulacra, areoles mostly circular but some elongated and overlapping with those on adjacent plates. A second series of primary tubercles—beginning near the interradius at the ambitus and remaining parallel to it (and therefore tracking slightly across the plate as the plates diminish in size adorally)—occurs fairly regularly on every second plate reaching to about two thirds to the peristome. This series may continue towards the peristome as secondary tubercles —in NIWA29524 tubercles in this series occurred on almost every plate in some columns. A third, shorter, series (but extending onto the aboral side) lies median to the other two occurring usually on every second plate and, with tubercles diminishing in size, can be traced about half way or slightly more down the column towards the peristome. Smaller secondary tubercles and granules (up to 15 or so) are scattered over the rest of the plates. A single series of primary tubs in each column of the ambulacra, parallel to and near the perradius, in some zones regularly on every third plate and in others less regular, on adjacent plates or every second plate. Plates otherwise covered with secondary tubercles and granules, 12 or so per plate.
Peristome 33 mm in diameter, bearing only ambulacral plates, boomerang-shaped, 11–12 per column. Each plate with 2 or 3 similar-sized tubercles on either side of the central tube-foot, their diameter about equal to the plate width, and a few granules bearing pedicellariae. Gills distinct, about 3mm long, with about 5 lobes. Buccal notches shallow.
Aboral test plating ( Figure 8 View FIGURE 8 ): In the interambulacra, the most prominent series of primary tubercles is the continuation of the median series of the oral side, occurring fairly regularly on every second plate and shifting gradually adradially as it is traced adapically, ending about two thirds up the column. A second series, comprising tubercles of widely varying sizes, occurs on most plates in the distal third to two-thirds of the column, forming a ragged line interradial to the primary series. This series not evident in some specimens examined. In a few specimens (e.g., NIWA45095), in some columns, two or three primary tubercles additionally found adradial to the primary series near the ambitus, occurring on alternating plates so as to form a discernable zig-zag pattern, but this is not the normal arrangement. Other smaller tubercles and granules (up to about 12) scattered over the rest of each plate. In the ambulacra, a series of tubercles of varying sizes (not exactly a continuation of the oral series but with similar irregularity) lies near the perradius and runs about two-thirds up the column adapically, on adjacent or every second or third plate. Secondary tubercles and granules (up to about 12) scattered over the rest of the plates. Accessory plates positioned midway along the primary plate and, although exact plate boundaries are obscured somewhat by skin and connective tissue, appear to be located quite separate from the primary plates—within the membrane between plates rather than incorporated within the primary plates. Tube-feet of primary plates located near their adradial edge, adapically.
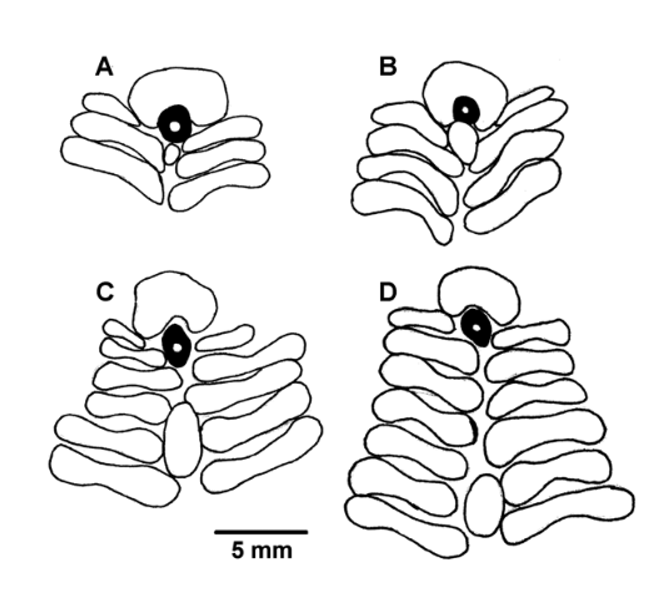
Apical system monocyclic, about 21 mm across, roughly pentagonal in outline. Exact plate structure difficult to determine accurately due to the covering of thick skin. Madreporite oval or kidney-shaped, only a little raised, pierced by 1 or 2 spines (their base being hidden below the surface), plate G2 enlarged slightly compared with the other genitals to accommodate it. Genital and ocular plates roughly triangular, the latter about a third the size of the former. Gonopores small, about 1 mm in diameter. Genital plates composite, a smaller distal plate separated from the larger plate by a membranous space pierced by the gonopore. These smaller plates evident in other specimens but not all, e.g. NIWA45095, in which the gonopores open into an elongated and pointed membranous space. However, in one interambulacral column of this specimen there is a larger additional plate located well beyond the genital pore, lying between about the 4th to the 6th plates from the apical system. This appears to be the distal component of the genital plate which has enlarged and migrated down the interradius. This unusual feature was also observed in several other specimens (NIWA45103, NIWA45994, NIWA45129, NIWA29415, NIWA53880, and NIWA23711), the enlarged plate sometimes reaching as far as the sixth or seventh interambulacral plate ( Figure 9 View FIGURE 9 ). Small tubercles and granules distributed evenly over several plates surrounding the anus, the anus itself surrounded by a fairly distinct ring of tubercles. Genital plates with about 15–16 small tubercles and granules, ocular plates with about 6–8. Shape of apical system variable, the genital pores opening into a more elongated membranous space in some specimens, giving the whole a more star-shaped appearance.
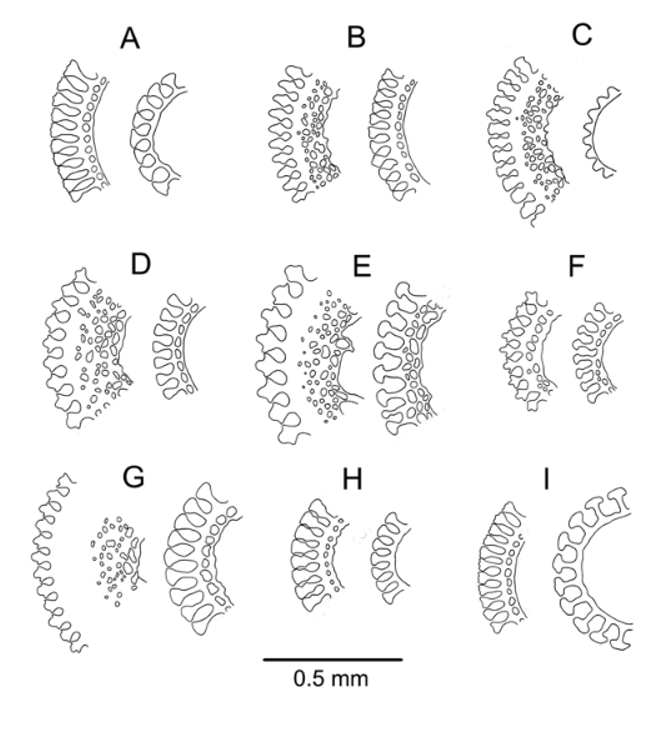
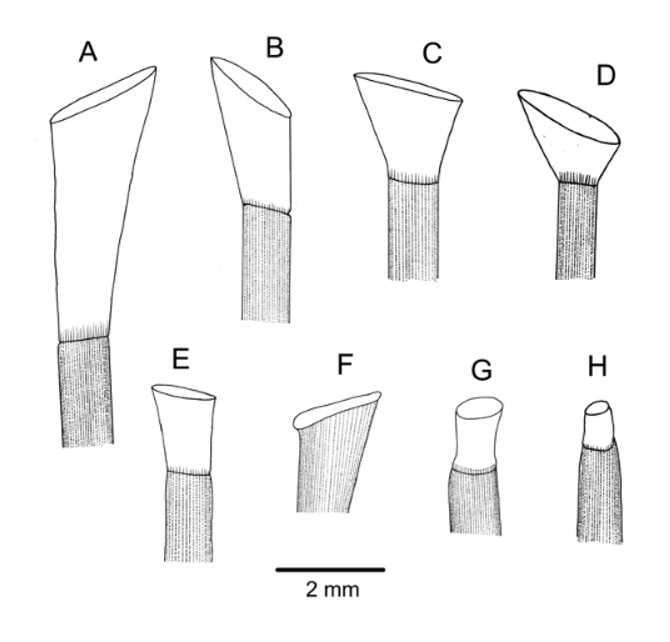
Spines: All primary spines broken near base, so lengths unknown, and no hoofs found. Intact spines on other specimens of similar size were up to 25 mm long and slightly curved. Some slightly damaged hoofs found on the oral spines of one specimen in NIWA53880 were white, short, and widely flaring ( Figure 4 View FIGURE 4 D). In addition, a single perfect hoof, white and almost hemispherical, was found on a specimen from the Tasman Sea NORFANZ survey (station TAN0308/134). Primary spines very delicate, circular, hollow, the largest about 0.7 mm diameter, smooth, with about 45 fine, longitudinal striations. These striations simple club-shaped wedges in cross-section in the oral spines examined, with an inner ring of perforations; those of the aboral spines formed of wedges with lateral extensions, outside of a thick layer of meshwork ( Figure 3 View FIGURE 3 A).
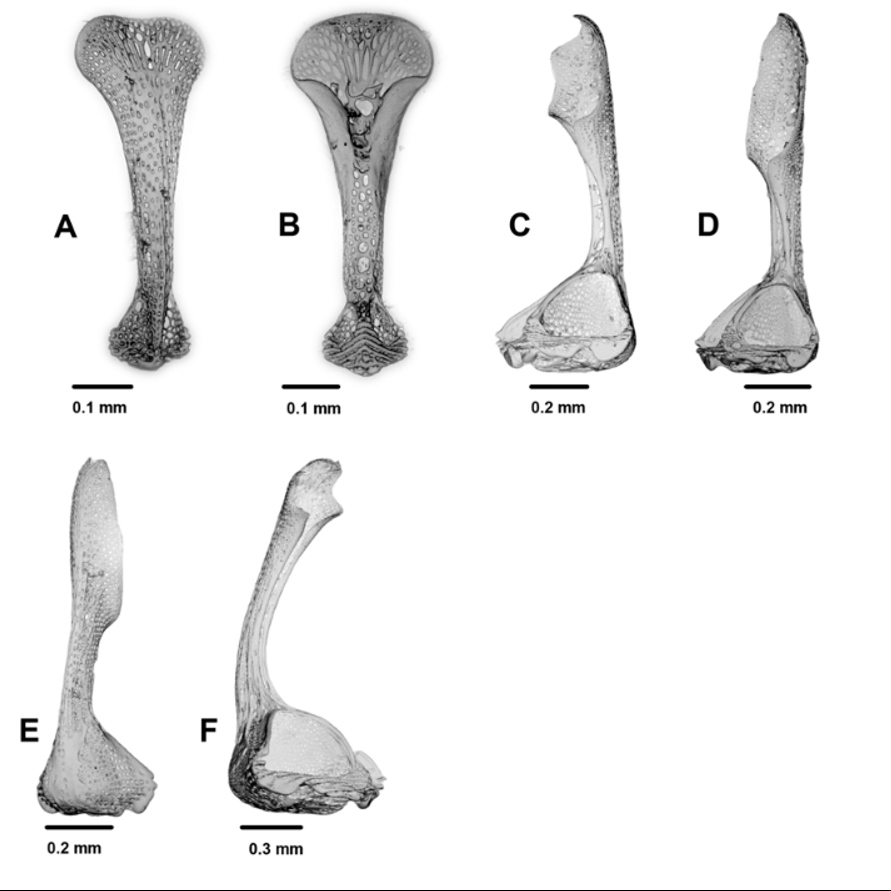
Pedicellariae ( Figure 10 View FIGURE 10 ): Three types of tridentate pedicellariae present: A large, distinctive, involute form with long, narrow, curved blades so strongly involuted that their edges almost touch along their entire length; head length about 1.7 mm, neck very short; blades widening only slightly to a very coarsely serrated tip, therefore providing only a small area of contact between them; the base large in proportion to the whole, allowing for significant muscle attachment. This form is quite common on the holotype and other specimens examined and is a distinctive feature of the species. Two smaller involute types also present, both with a head length of about 1.0– 1.2 mm. One with rounded, coarsely serrated blade tips in contact over about a third of the valve length, a relatively large base, and a neck about half the valve length; the valve strongly involuted proximal to the blade. The other with narrow, un-serrated blade tips in contact for nearly half the valve length, a relatively small base, and a neck about two-thirds the valve length. The blade of this type also strongly involuted proximal to the blade. Triphyllous pedicellariae: long and narrow (head length about 0.5 mm); blades evenly curved laterally, with a rounded tip; basal section of blades involuted, the edges joined along much of their length to form a hollow scoop; blade width nearly twice base width; neck long (about 2–4 times head length), stalk up to 10 times head length. Dactylous forms also present but very rare and not found on the holotype; a single, broken specimen found on NIWA29415 (with 4 valves) and one on NIWA45994 (with 5 valves).
Sphaeridia; elongated, club or sausage-shaped (about 1.0 mm long by 0.2 mm wide), transparent, located adjacent (slightly perradial and adoral) to the inner tube foot of the ambulacral plates.
One specimen from about 600 m on the Campbell Plateau (NIWA46090) is unusual in that it is very large (178 mm TD) with a tough, reddish-pink test distinctly raised aborally so that in general appearance it quite strongly resembles A. alternatum (see below), but the continuous series of oral interambulacral adradial tubercles clearly shows that it is not this species. Although the inner series of tubercles in these same columns differ from the usual A. migratum arrangement in that they occur on nearly every plate in most columns, the pedicellariae and presence of displaced genital plates in the interambulacra are strongly characteristic of the species. Although included here under A. migratum this is nevertheless an unusual example and may possibly be a hybrid with the larger A. alternatum as both species occur in this area and depth range.
Size range —The median test diameter of the 17 specimens measured was 134 mm TD, and the largest specimen (excluding the unusual NIWA46090) was 166 mm TD.
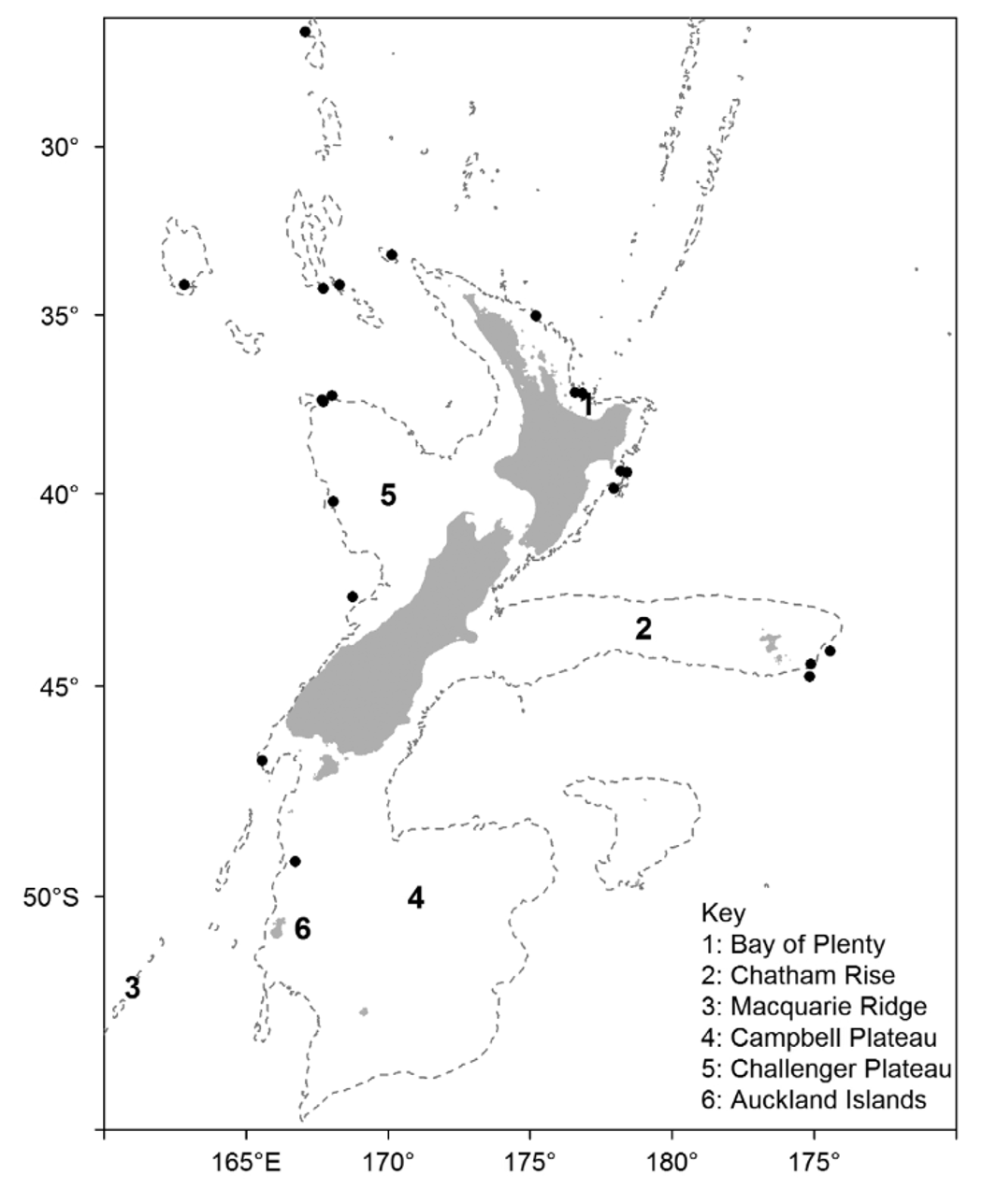
Occurrence —There have been 25 confirmed records of A. migratum since the first specimen was taken in 1967, representing 47 individuals.
These records are spread widely over the New Zealand region as well as south-eastern Australia ( Figure 11 View FIGURE 11 ) and virtually all records are in the vicinity of seamounts and other undersea features where trawling for deepsea fish such as orange roughy, alfonsino, and oreos takes place. Most of the New Zealand records were collected either by government fisheries observers working on board vessels in these fisheries, or by scientists on RV Tangaroa studying these fisheries or their effects on seamount ecosystems. The most northern records (from the Lord Howe Rise, Norfolk Ridge, and West Norfolk Ridge) were mostly collected by this vessel during the 2003 NORFANZ survey. The easternmost records come from the eastern end of the Chatham Rise and the most southern records from north of the Auckland Islands on the Campbell Plateau. Taking into account the records scattered off the west and east coasts of New Zealand it seems likely that the species is widespread over the entire region in suitable habitat. The records from Tasmania confirm its existence also on the western side of the Tasman Sea and so it also may be present over a wide area around southeast Australia.
The conservative depth range for the species is 614–949 m, with a potential range of 593–1170 m. The average difference between the start and finish depths for these trawls was 115 m, not unusual when bottom trawling on seamounts, the conservative depth range therefore being more reliable in this case.
Remarks —The species most similar to A. migratum seem to be A. leptaleum (known from Panama (Mortensen 1935) and the Galapagos (Pawson & Ahearn 2001)) and A. fenestratum (known from the North Atlantic, where it is widespread, and the Caribbean (Clark 1941)).
A specimen of A. fenestratum from off the Florida coast in the North Atlantic examined in the collection of Museum Victoria was very similar in colour, test plate morphology, and tuberculation to A. migratum , and also had similarly broad membranous spaces between the coronal plates. A. migratum and A. fenestratum also share a similar set of pedicellariae, but these differ in a fairly constant manner. The equivalent large tridentate form in A. fenestratum has a much shorter involute section and this section and the blade end are relatively wider than in A.
migratum . The A. migratum form of involute tridentate pedicellaria is also found in A. bakeri sp. nov. and Calveriosoma gracile , species which are otherwise well differentiated from A. migratum by their test morphology and colour. The two smaller forms of tridentate pedicellariae in A. migratum also have approximate counterparts in A. fenestratum , but these too differ conspicuously in the length of the involuted section of the blade (see figures in Thomson 1874 and Mortensen 1903 for comparison). The triphyllous and dactylous pedicellariae, the least variable forms within the Echinothuriinae , are as far as can be ascertained fairly similar between the two species. Finally, the hoofs on the oral spines appear to be shorter and broader in A. migratum .
The similarity between A. migratum and A. leptaleum is also apparent in the morphology and tuberculation of the coronal plates. However, A. leptaleum has distinctive, tapering hoofs, more conspicuously broad membranous spaces between the oral interambulacral plates, and a poorer variety of pedicellariae (Agassiz & Clark 1909). The specimen of A. leptaleum figured by Agassiz & Clark (1909), and their description, indicate a compound genital plate as described for A. migratum , but there is no suggestion that the distal plate becomes separated from the others as it frequently does in this species. In fact, the unusual migration of genital plates in A. migratum has not been reported in any other echinothurioid.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.