
Glycera nicobarica Grube, 1866
|
publication ID |
https://doi.org/ 10.12782/sd.18.2.269 |
|
persistent identifier |
https://treatment.plazi.org/id/2C08878C-FF87-FFF9-16DE-FA00D77CCCCD |
|
treatment provided by |
Felipe |
|
scientific name |
Glycera nicobarica Grube, 1866 |
| status |
|
Glycera nicobarica Grube, 1866 View in CoL
[Japanese name: Chirori] ( Figs 1–9 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig )
Glycera nicobarica Grube, 1866: 178 View in CoL ; Grube 1867: 24–25, pl. 3, figs 1, 1a, b; Böggemann 2002: 57–58, 124–125, figs 67–69; Imajima 2003: 112–113, fig. 67 a–h; Imajima 2007: 30–231, fig. 71.
Glycera decipiens Marenzeller, 1879: 140 View in CoL , pl. 6, figs 3, 3A.
Glycera amboinensis McIntosh, 1885: 345–346 View in CoL , pl. 42, figs 6, 7.
Glycera chirori Izuka, 1912: 245–246 View in CoL , pl. 2, fig. 8, pl. 24, fig. 13; Okuda 1938: 125, fig. 2; Imajima and Hartman 1964: 161–162; Uchida 1992: 319.
Glycera hasidatensis Izuka, 1912: 246–247 View in CoL , pl. 24, figs 14, 15 (in part).
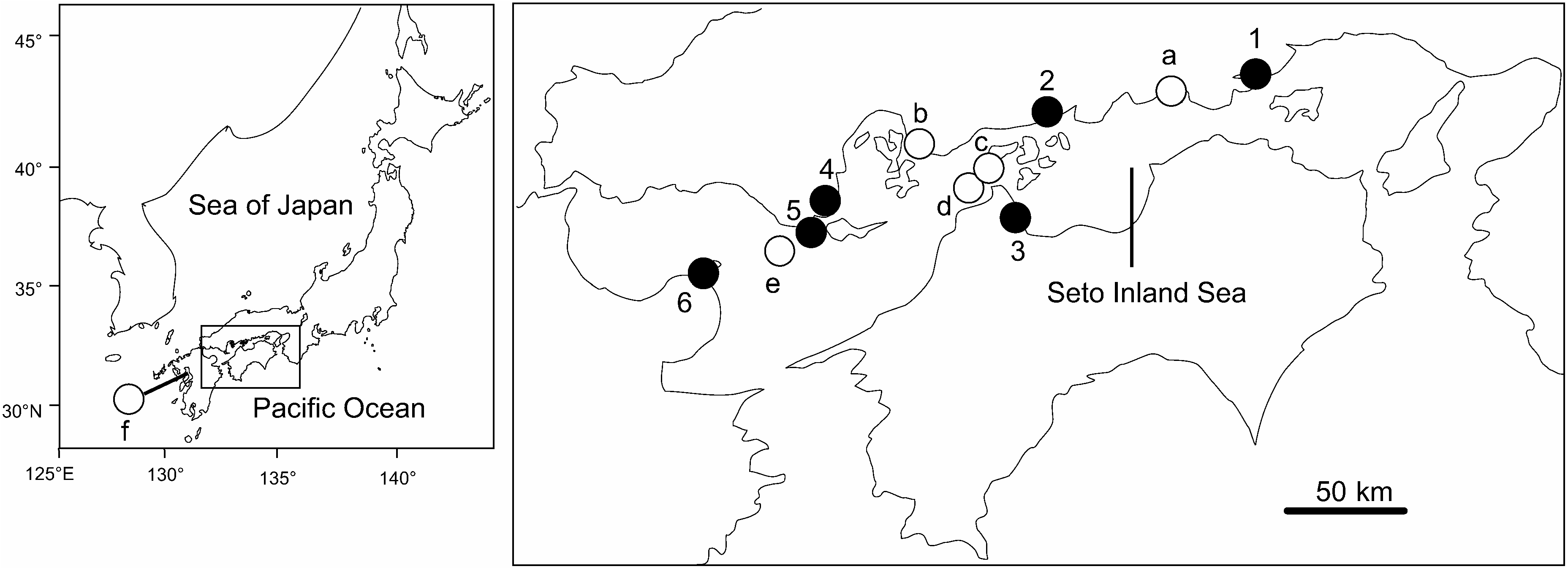
Material examined. Sexually mature specimens with epitokous metamorphosis (epitokes) in the Seto Inland Sea: Uno Port (34°29′33″N, 133°57′26″E, 12 m deep), Okayama Prefecture, 9–11 November 2010, coll GoogleMaps . T. Urata, 10 specimens (MS); 8–10 November 2011, coll . Y . S . Ibrahim, 33 specimens (NSMT-Pol 111422–111423, OMNH-Iv 5354– 5355, 5357–5358, SMF 22825–22827 About SMF , MS) . Imabari Port (34°4′6″N, 133°0′29″E, 7.4 m deep), Ehime Prefecture, 27 October 2009, coll GoogleMaps . M GoogleMaps . Sato et al., 14 specimens (CMNH- ZW 002074–002075 , MS) . Himeshima Port (33°43′17″N, 131°38′47″E), Oita Prefecture, 28 October 2009, coll GoogleMaps . M GoogleMaps . Sato et al., 11 specimens (MS). Yanai Port (33°57′24″N, 132°8′10″E), Yamaguchi Prefecture, 29 October 2009, coll GoogleMaps . M GoogleMaps . Sato et al., 9 specimens (NSMT-Pol 111427, MS); 9 July 2010, coll . Y GoogleMaps . S. Ibrahim and M . Sato, 2 specimens (NSMT- Pol 111426, MS) . Tadanoumi (34°19′57″N, 132°59′8″E), Takehara , Hiroshima Prefecture, 10 July 2010, coll GoogleMaps . Y GoogleMaps . S. Ibrahim and M . Sato, 5 specimens (NSMT-Pol 111424– 111425, CMNH-ZW 002077–002078 , MS) . Off Tanoura (33°47′27″N, 132°1′59″E), Nagashima Island, Kaminoseki , Yamaguchi Prefecture, 30 July 2011, coll GoogleMaps . Y GoogleMaps . S. Ibrahim, 4 specimens (MS).
Immature specimens (atokes) in the Seto Inland Sea: Intertidal flats in Kasaoka Bay (34°27′–29′N, 133°31′– 32′E), Okayama Prefecture, 13 October 2004, coll . K . Okada, 1 specimen (NSMT-Pol 111428). Intertidal flats in Kawajiri (34°13′1″N, 132°40′13″E), Kure , Hiroshima Prefecture, 26 February 1998, coll GoogleMaps . M GoogleMaps . Sato, 5 specimens (NSMT- Pol 111430, MS) . Bouchi-no-su (34°22′29″–22′38″N, 133°36′52″–36′53″E, 23–24 m deep), north of Sanagishima Island in the Shiwaku Islands, Kagawa Prefecture, 8 November 2011, coll. Y. S. Ibrahim, 20 specimens (OMNH-Iv 5356, CMNH-ZW 002076 , SMF 22828, MS). Nukari-no-seto (34°18′29″N, 133°13′41″E, 14–15 m deep), Hiroshima Prefecture, 8 November 2011, coll. Y. S. Ibrahim, 3 specimens (MS). Off Iwaijima Island (33°47′18″–47′52″N, 131°59′11″– 59′22″E, 20–27 m deep), Yamaguchi Prefecture, 6 May 2000, coll GoogleMaps . H GoogleMaps . Fukuda, 6 specimens (MS).
Atoke in Ariake Sea: Kojiro-nagahama (32°52′20″N, 130°15′41″E), Isahaya Bay , Kunimi Town , Nagasaki Prefecture, 2 September 2003, coll GoogleMaps . M GoogleMaps . Sato, 1 specimen (NSMT- Pol 111429).
Diagnosis (based on Böggemann 2002). Two types of papillae present on proboscis: conical papillae with 3 Ushaped transverse ridges and oval ones without transverse ridges. Ailerons with triangular bases. Biramous parapodia with two prechaetal lobes (notopodial and neuropodial lobes slender, digitiform, of same length throughout) and two postchaetal lobes (notopodial lobe triangular; neuropodial lobe shorter and rounded). Simple digitiform branchiae, situated medially on anterior side of parapodia.
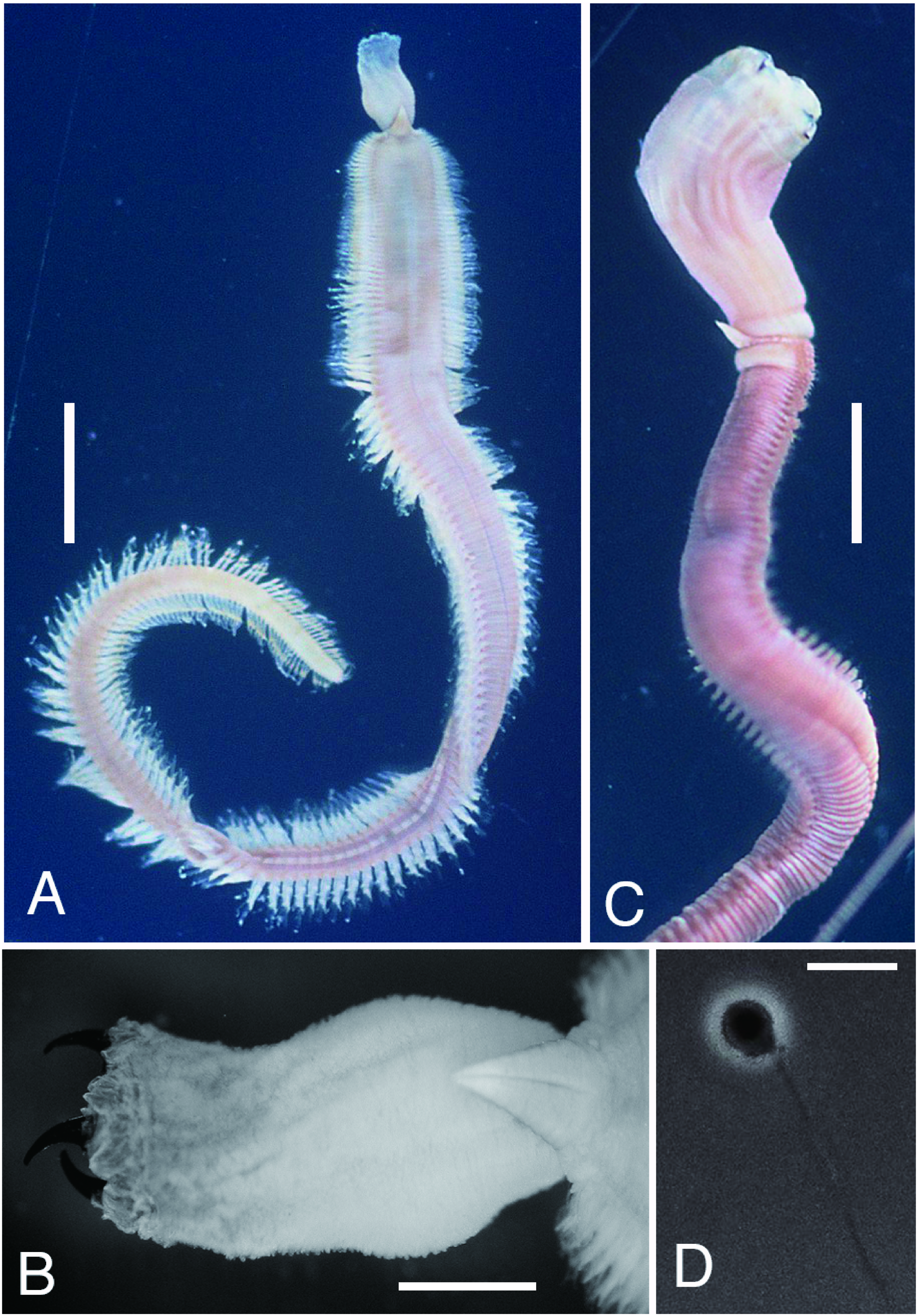
Description of epitokes [based on 88 (18 complete and 70 incomplete) specimens]. Body flattened, 0.8–3.6 mm in BW, 0.4–2.7 mm in BH ( Fig. 2A View Fig ). Largest complete specimen (male) 96 mm in BL, 2.8 mm in BW, with 218 chaetigers. Smallest complete specimen (spent worm) 23 mm in BL, 1.6 mm in BW, with 135 chaetigers. Body color in preserved specimens pale brown to dark brown in both males ( Fig. 2A View Fig ) and females. No significant difference in BW between males (range 1.8–3.6 mm, mean±SD 2.5± 0.5 mm, n =21) and females (2.0– 3.2 mm, 2.5± 0.5 mm, n =4) (Wilcoxon-Mann-Whitney test, P =0.88). Sex undeterminable for 63 spent worms (0.8–3.2 mm in BW) with all gametes discharged before collection.
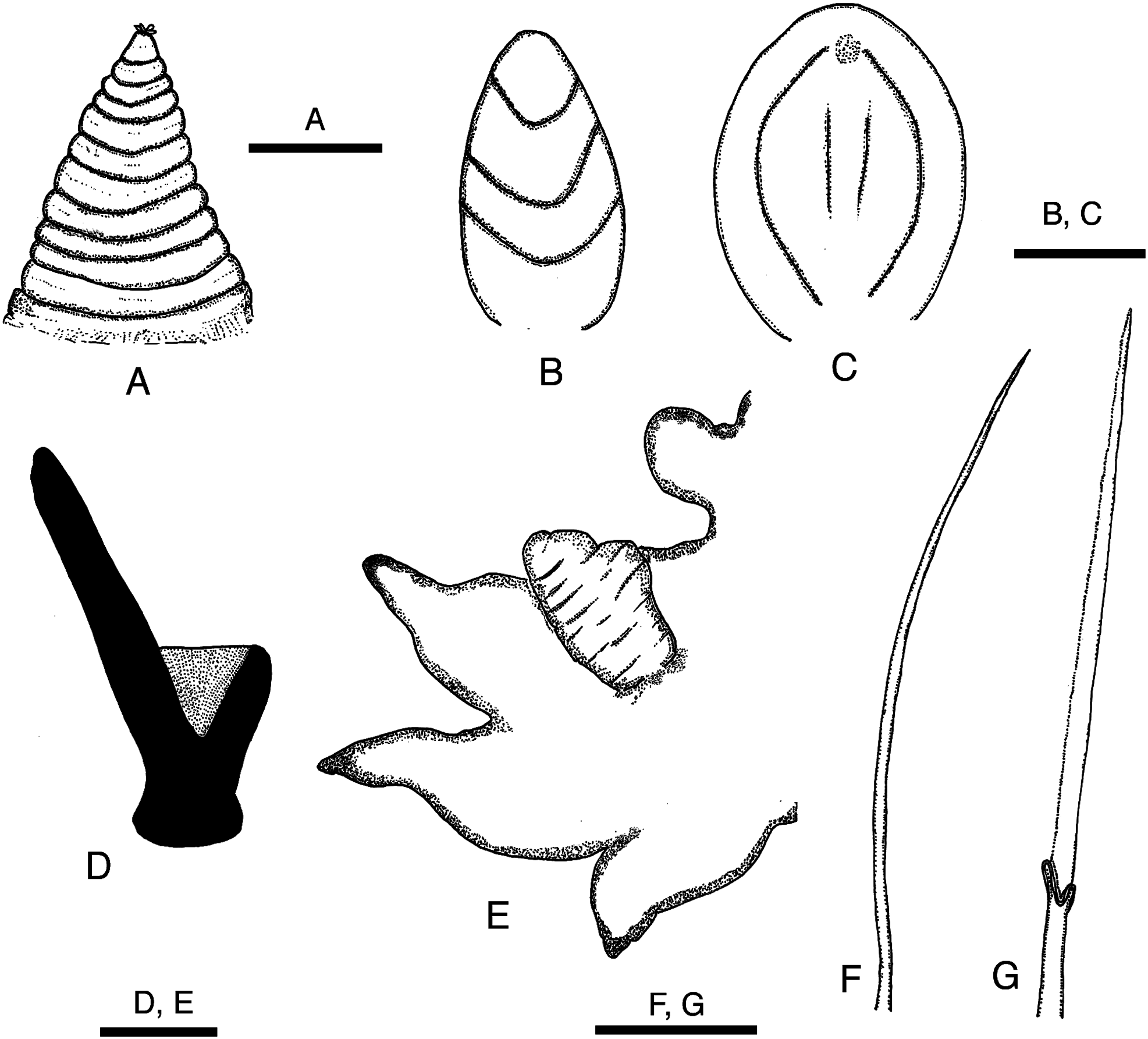
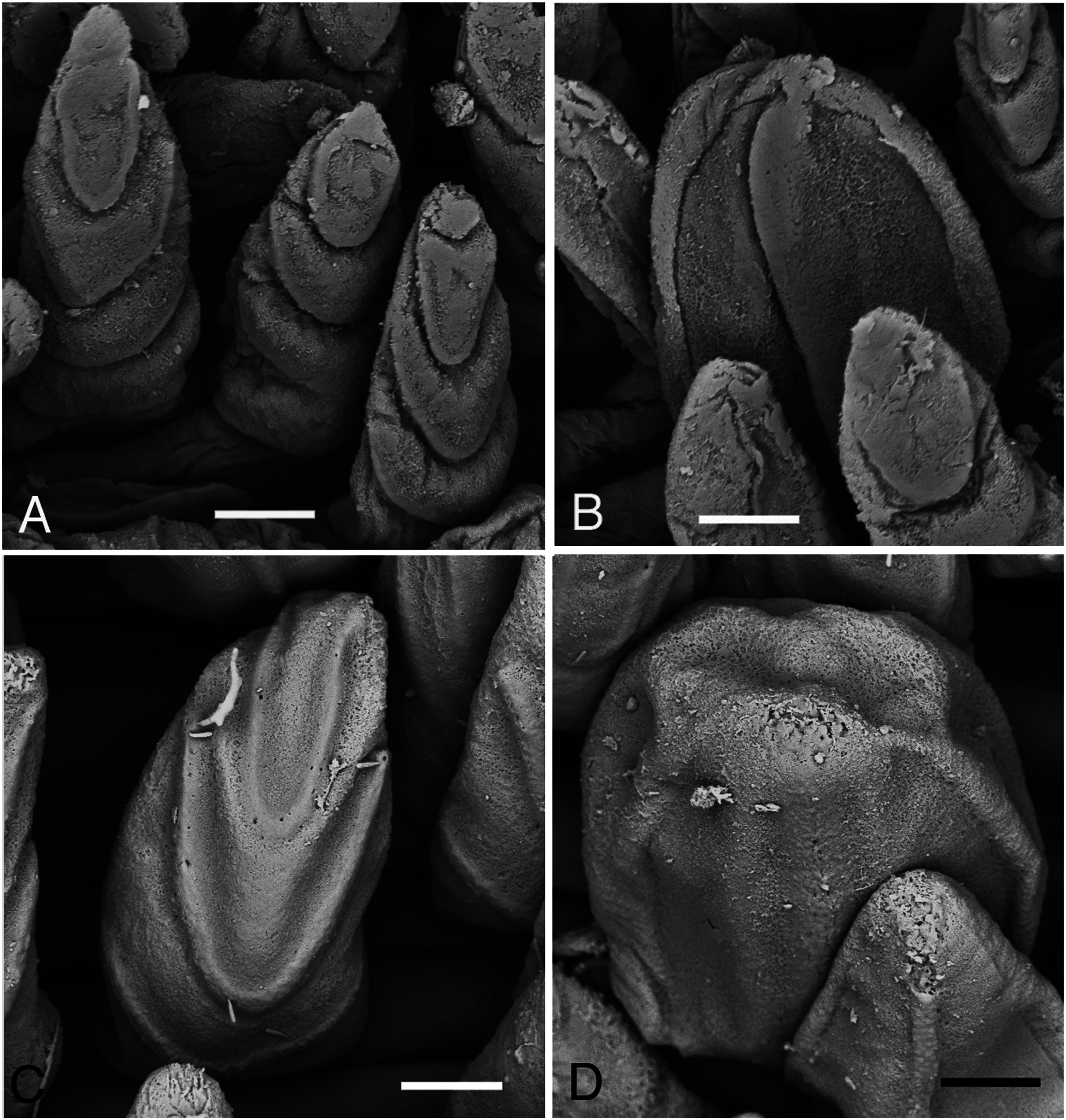
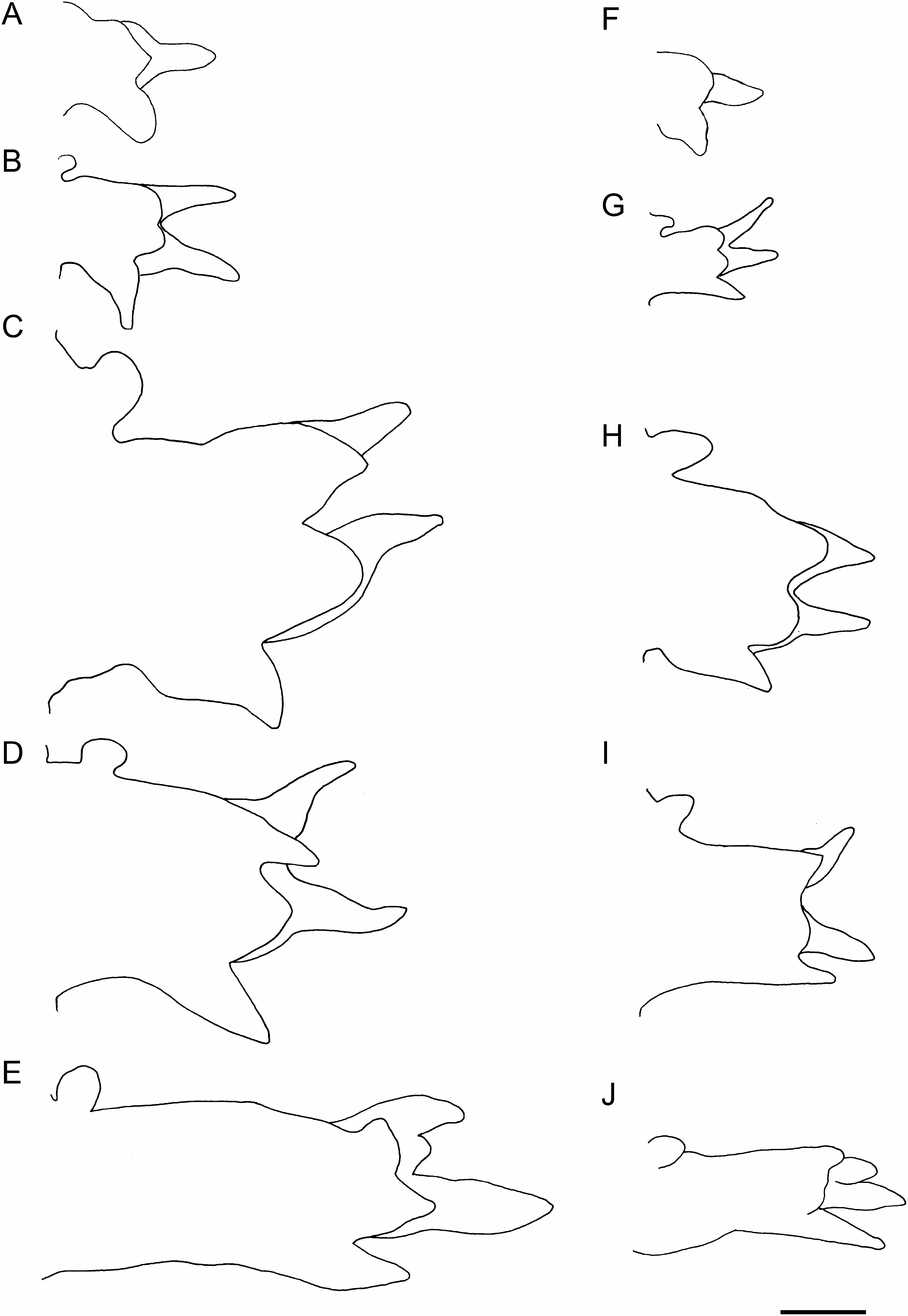
Prostomium conical, consisting of 9 to 11 rings altogether as long as first 11–13 chaetigers ( Figs 2A, B View Fig , 3A View Fig ). Proboscis cylindrical, PL 4–10mm (7–23% of BL), as long as first 20–40 chaetigers, PW 1–2 mm ( Fig. 2A, B View Fig ). Two types of papillae present on proboscis: conical ones with 3 U-shaped ridges ( Figs 3B View Fig , 4A View Fig ) and oval ones without ridges ( Figs 3C View Fig , 4B View Fig ). Four black, hook-like jaws present on tip of proboscis ( Fig. 2B View Fig ); aileron with triangular base ( Fig. 3D View Fig ) present at base of each jaw.
First 2 pairs of parapodia uniramous, with single acicula, digitiform prechaetal lobe, shorter, rounded postchaetal lobe, and thick, rounded ventral cirrus; ventral cirrus longer than postchaetal lobe ( Fig. 5A View Fig ). All following parapodia biramous, with 2 aciculae, 2 prechaetal lobes, 2 postchaetal lobes, and ventral cirrus. Two prechaetal lobes slender and triangular, of almost same length in anterior and middle parapodia; neuropodial prechaetal lobe longer than notopodial one in posterior parapodia. Two postchaetal lobes shorter than prechaetal lobes throughout; both lobes round- ed, of same length in anteriormost parapodia back to about chaetiger 10; notopodial postchaetal lobe triangular with tapering tip, slightly longer than round neuropodial one in middle parapodia ( Fig. 5B–D View Fig ); both lobes triangular, of same length in posterior parapodia ( Fig. 5E View Fig ). Dorsal cirrus absent in parapodium 1, usually present from parapodium 2 (sometimes from parapodium 3) onward; dorsal cirrus oval, inserted into body wall above parapodial base in anterior body back to about chaetiger 20; dorsal cirrus attached to upper edge of parapodial base in following chaetigers. Ventral cirrus triangular, about as long as neuropodial postchaetal lobe throughout.
Branchiae retractile, simple, and digitiform, appearing from parapodia 16–31 to near posterior end, situated medially on anterior side of parapodia, largest in middle or posterior parapodia ( Fig. 3E View Fig ).
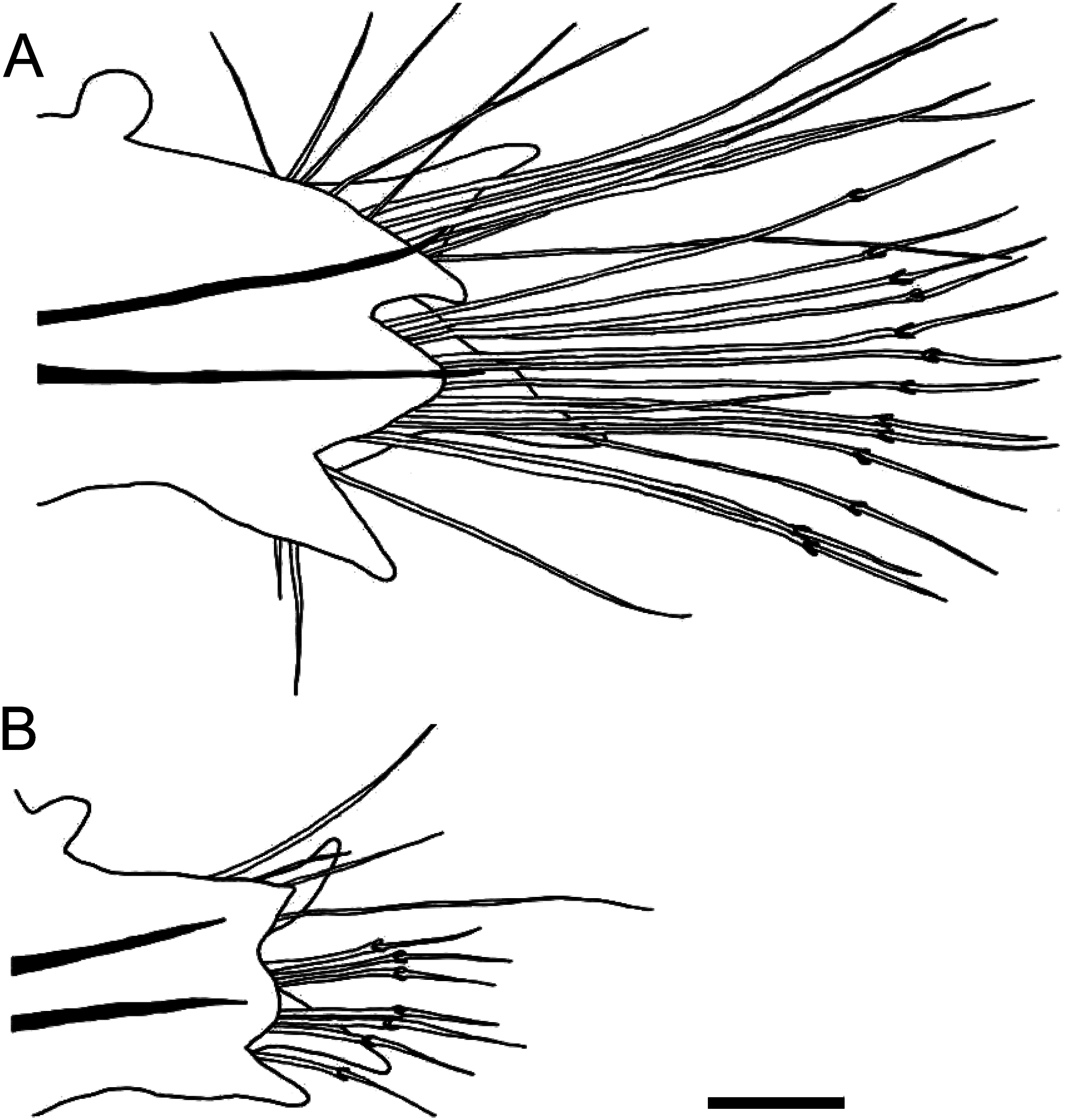
Notochaetae all simple capillaries throughout, 3–13, 4–19, and 2–8 in anterior (chaetigers 10–20), middle (chaetigers 50–110), and posterior (chaetigers 160–210) parapodia, respectively ( Figs 3F View Fig , 6A View Fig ). Upper neurochaetae all spinigerous compound chaetae, numbering 1–11, 1–11, and 1–4 in anterior, middle, and posterior parapodia, respectively ( Figs 3G View Fig , 6A View Fig ). Lower neurochaetae consisting of spinigerous compound chaetae at upper position, numbering 0–17, 2–16, and 1–8 in anterior, middle, and posterior parapodia, respectively; simple capillaries at lower position, numbering 0–3, 0–7, and 0–2 in anterior, middle, and posterior parapodia, respectively. All chaetae elongated, longest in middle parapodia.
Head of spermatozoa round, about 3 µm long, 3 µm wide ( Fig. 2D View Fig ). Oocytes 140–150 µm in diameter.
Comparison of morphology between atokes and epitokes. Morphological characteristics of the 88 swimming epitokes (0.8–3.6 mm in BW, mean±SD: 2.1± 0.5 mm) were compared with those of 35 benthic atokes of comparative body size (BW 1.0–3.0 mm, 2.2± 0.5 mm); there was no significant difference in BW between the epitokes and atokes (Wilcoxon-Mann-Whitney test, P =0.07). Epitokous males and females were pale or dark brown in body color whereas both living and ethanol-preserved specimens of atokes were reddish-brown ( Fig. 2A, C View Fig ). The body wall of epitokes was thinner than that of atokes.
The body became flattened in epitokes. BH of epitokes (0.4–2.7 mm, 1.2± 0.5 mm, n =88) was significantly less than that of atokes (0.9–3.0 mm, 2.0± 0.5 mm, n =35) (P <0.0001). The thickness index (BH/BW) of epitokes (0.2– 1.0, 0.6±0.2, n =88) was significantly smaller than that of atokes (0.9–1.0, 0.9±1.1, n =35) (P <0.0001).
The proboscis of epitokes was markedly reduced in size. PL of epitokes (3.2–5.4 mm, 4.0± 0.8 mm, n =10) was significantly smaller than that of atokes (3.3–9.8 mm, 6.0± 2.0 mm, n =14) (P =0.005) ( Fig. 2A, C View Fig ). PW of epitokes (1.0– 1.7 mm, 1.4± 0.2 mm, n =10) was significantly small- er than that of atokes (2.1–4.5 mm, 3.0± 0.7 mm, n =14) (P <0.0001).
The parapodia of epitokes ( Fig. 5A–E View Fig ) were enlarged throughout in comparison to those of atokes ( Fig. 5F–J View Fig ), with chaetae elongated and increased in number ( Fig. 6A, B View Fig ). Numbers of notochaetae (simple capillaries, up to 12), upper neurochaetae (spinigerous compound chaetae, up to 8), and lower neurochaetae (spinigerous compound chaetae, up to 11) in atokes (n =3) were smaller than those of epitokes (up to 19, 11, and 17, respectively) (n =13). Simple capillaries appeared not only on notopodia but also on neuropodia (inferior portion) in epitokes, whereas they appeared only on notopodia in atokes.
Symbionts. Several small organisms including copepods ( Fig. 7A, B View Fig ) and nematodes ( Fig. 7C, D View Fig ) were attached to the body wall and parapodia of four epitokous males and seven spent worms ( Fig. 7 View Fig ). The copepods ( Fig. 7A, B View Fig ) were identified as belonging to the family Clausiidae (Copepoda: Cyclopoida ) (Susumu Ohtsuka, pers. comm.). The relationship between these symbionts and the epitokous worms is unknown. No symbionts were observed in atokous specimens.
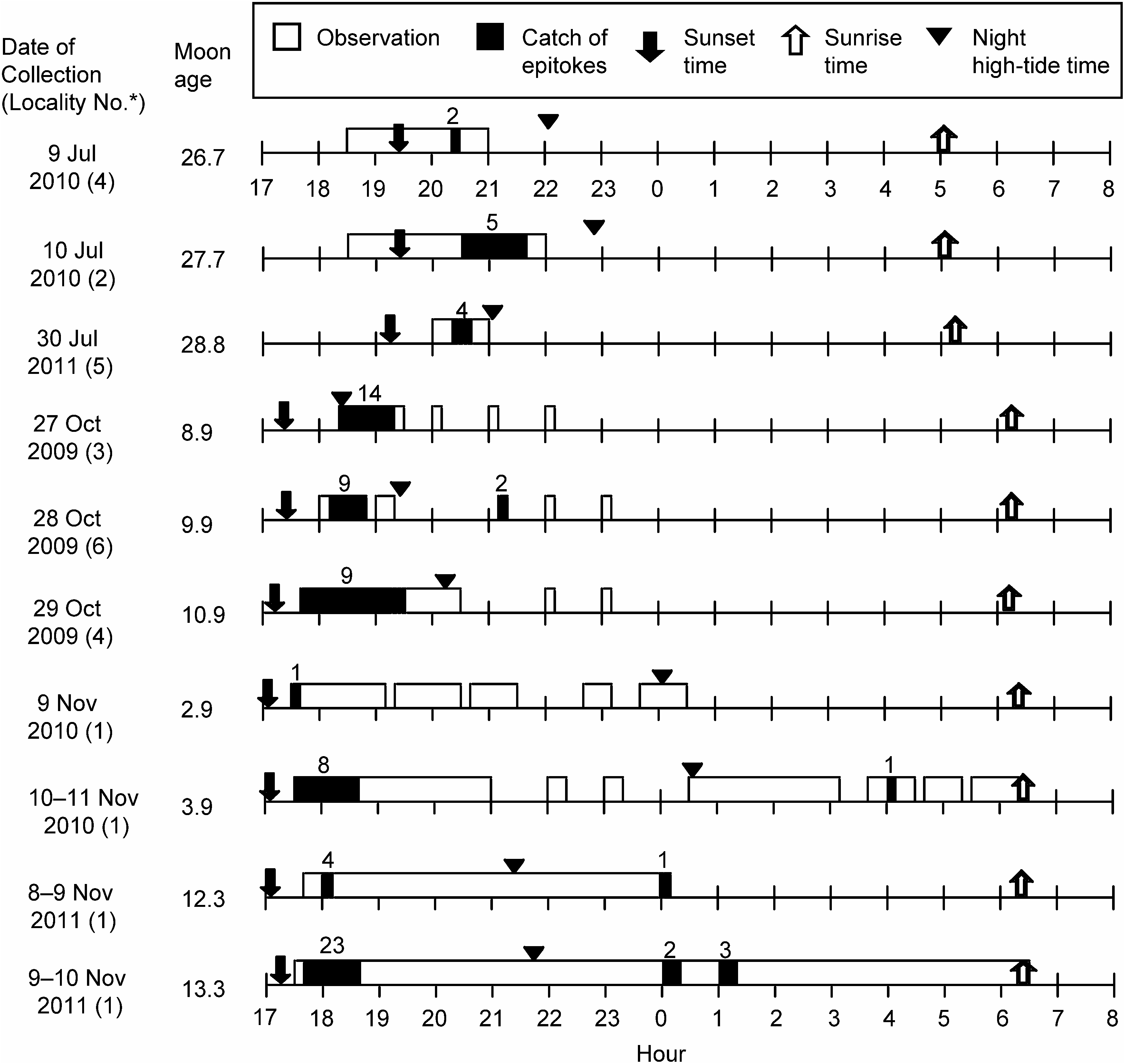
Swimming behaviour of epitokes. Epitokes were collected throughout our sampling period (July to November) in the Seto Inland Sea. Many specimens were collected from the end of October to early November ( Fig. 8 View Fig ). Their dates of occurrence were not related to any specific moon phases (2.9–28.8). Water temperatures and salinities at sampling sites were recorded as 20–25°C and 29–32 psu, respectively. Most epitokes (n =81) were collected in the evening (17:00– 21:40) within two hours after sunset, although a few epitokes were collected at midnight (0:00–1:15, n =6) or early morning before sunrise (4:05, n =1), with no relation to tidal condition ( Fig. 8 View Fig ).
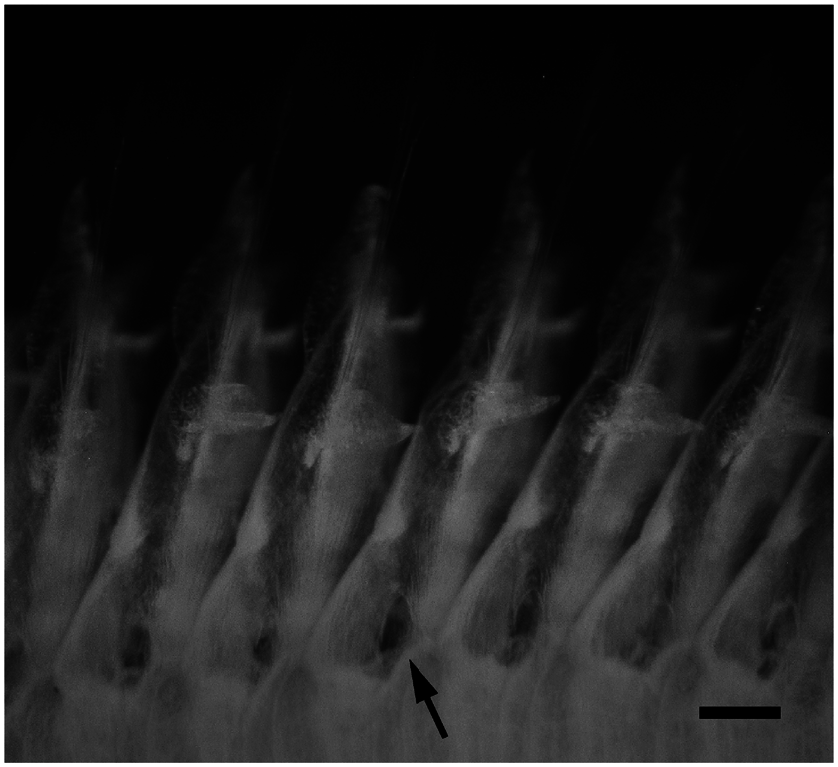
Epitokes swam slowly, straight and head first in surface water, showing a wave-like movement. We do not use the term “swarming” for their swimming behaviour because we did not find mass swimming, which is well-known in epitokes of some nereidids, syllids, and eunicids ( Clark 1961). In most cases, we observed only a few individuals swimming at the surface water except in Uno port on 9 November 2011, when we collected relatively many epitokes within a short period: 17 individuals in 15 min (18:15–18:30) ( Fig. 8 View Fig ). We observed some epitokes shedding their whitish gametes through the body wall during swimming. Small holes or slits, which are probably the sites of gamete discharge, were often found around the ventral surface of the base of the parapodia in the middle body (chaetigers 47–143) of 22 epitokous worms ( Fig. 9 View Fig ).
Geographical distribution. Indian Ocean, coasts of Japan, East and South China Seas ( Böggemann 2002).
Remarks. The author and year of the original description of Glycera nicobarica has been referred to as “Grube, 1868” by previous authors (e.g., Böggemann 2002). But recently Böggemann (pers. comm.) discovered an earlier publication of the description of this species ( Grube 1866).

| T |
Tavera, Department of Geology and Geophysics |
| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Glycera nicobarica Grube, 1866
| Ibrahim, Yusof Shuaib & Sato, Masanori 2013 |
Glycera chirori
| Uchida, H. 1992: 319 |
| Imajima, M. & Hartman, O. 1964: 161 |
| Okuda, S. 1938: 125 |
| Izuka, A. 1912: 246 |
Glycera hasidatensis
| Izuka, A. 1912: 247 |
Glycera amboinensis
| McIntosh, W. C. 1885: 346 |
Glycera decipiens
| Marenzeller, E. 1879: 140 |
Glycera nicobarica
| Imajima, M. 2007: 30 |
| Imajima, M. 2003: 112 |
| Boggemann, M. 2002: 57 |
| Grube, E. 1867: 24 |
| Grube, E. 1866: 178 |