
Hemienchytraeus tenuiculus, Chen & Schmelz & Zhang & Xie, 2022
|
publication ID |
https://doi.org/ 10.1080/00222933.2022.2140085 |
|
DOI |
https://doi.org/10.5281/zenodo.7426642 |
|
persistent identifier |
https://treatment.plazi.org/id/3A40CF59-FF94-FFC2-B2B2-43E3FB93DA51 |
|
treatment provided by |
Plazi |
|
scientific name |
Hemienchytraeus tenuiculus |
| status |
sp. nov. |
Hemienchytraeus tenuiculus sp. nov.
( Figures 6–8 View Figure 6 View Figure 7 View Figure 8 )
Holotype
Fully mature, whole-mounted specimen, stained, GZO202007012 .
Type locality
Site 2. Soil and moss layer of Cyclobalanopsis glauca forest (108°42′58.7″E, 27°54′28.91″N), 2000 m asl, Fanjing Mountain , Guizhou, China, coll GoogleMaps . Z GoogleMaps .X. Zhang, Y.H. Ge and J.J. Chen, 25 December 2019.
Paratypes
GZO202007013, GZO20201010001 , stained and whole-mounted, two mature specimens from the type locality, same data as holotype . CJJ81 , CJJ125 , CJJ126 , CJJ130 , four mature specimens from site 2, CJJ 82 , one mature specimen from site 4, CJJ 83 , one mature specimen from site I, CJJ 132 , one mature specimen from site 8: seven specimens in total, whole worm used for DNA extraction, preserved as total DNA .
Further material investigated
One mature specimen from site 6, one mature specimen from site 10 and one mature specimen from site 12: three specimens in total, preserved in 75% ethanol.
Etymology
Named for the small and thin body.
Diagnosis
The species can be diagnosed by the following combination of characters: (1) worms small and thin; (2) each primary branch bifurcating into two secondary branches; (3) spermathecae extending to VII–VIII; (4) no secondary pharyngeal glands; (5) three pairs of preclitellar nephridia in 6/7–8/9; (6) dorsal vessel originating from postclitellar segments; (7) clitellum girdle-shaped.
Description
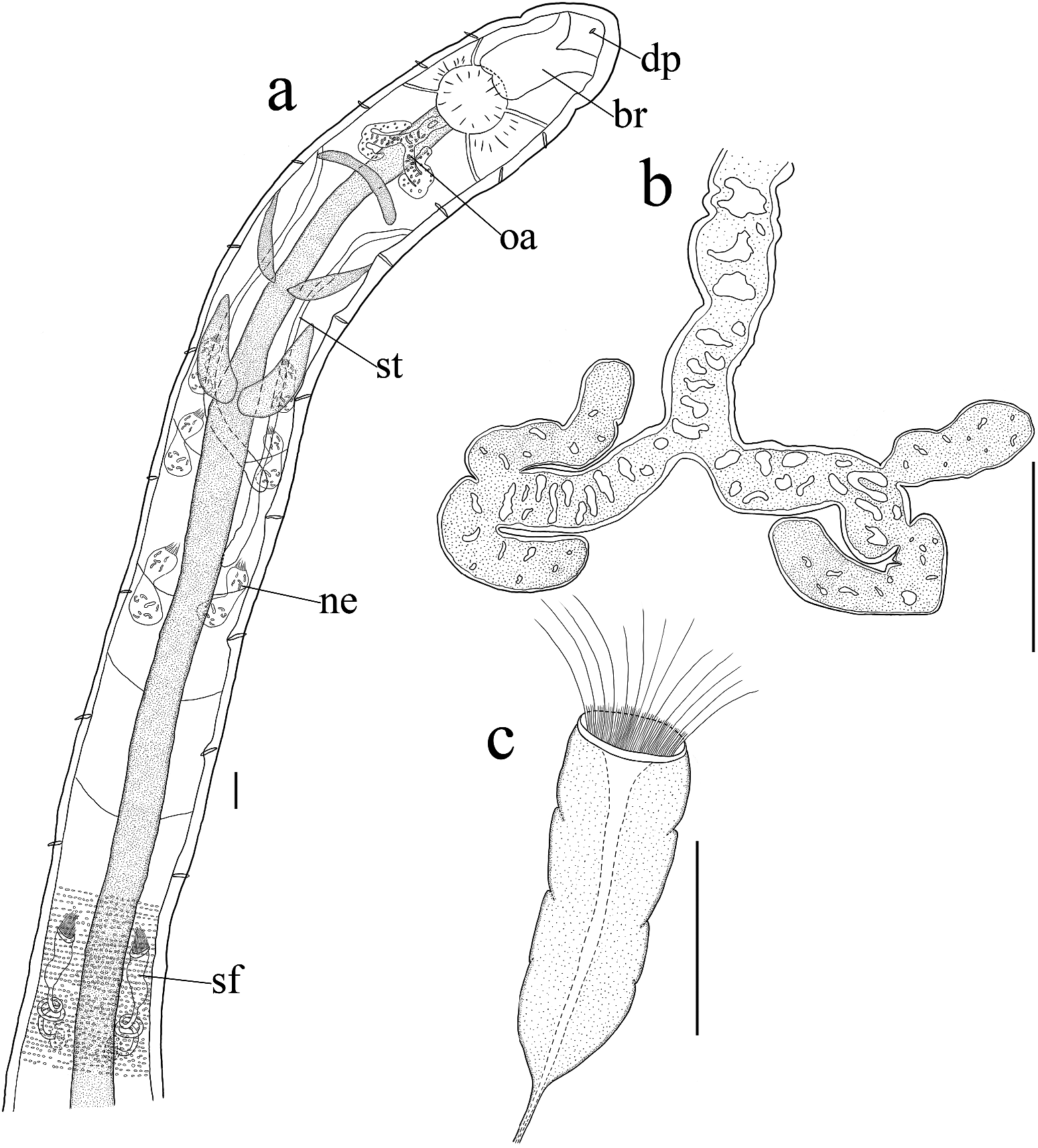
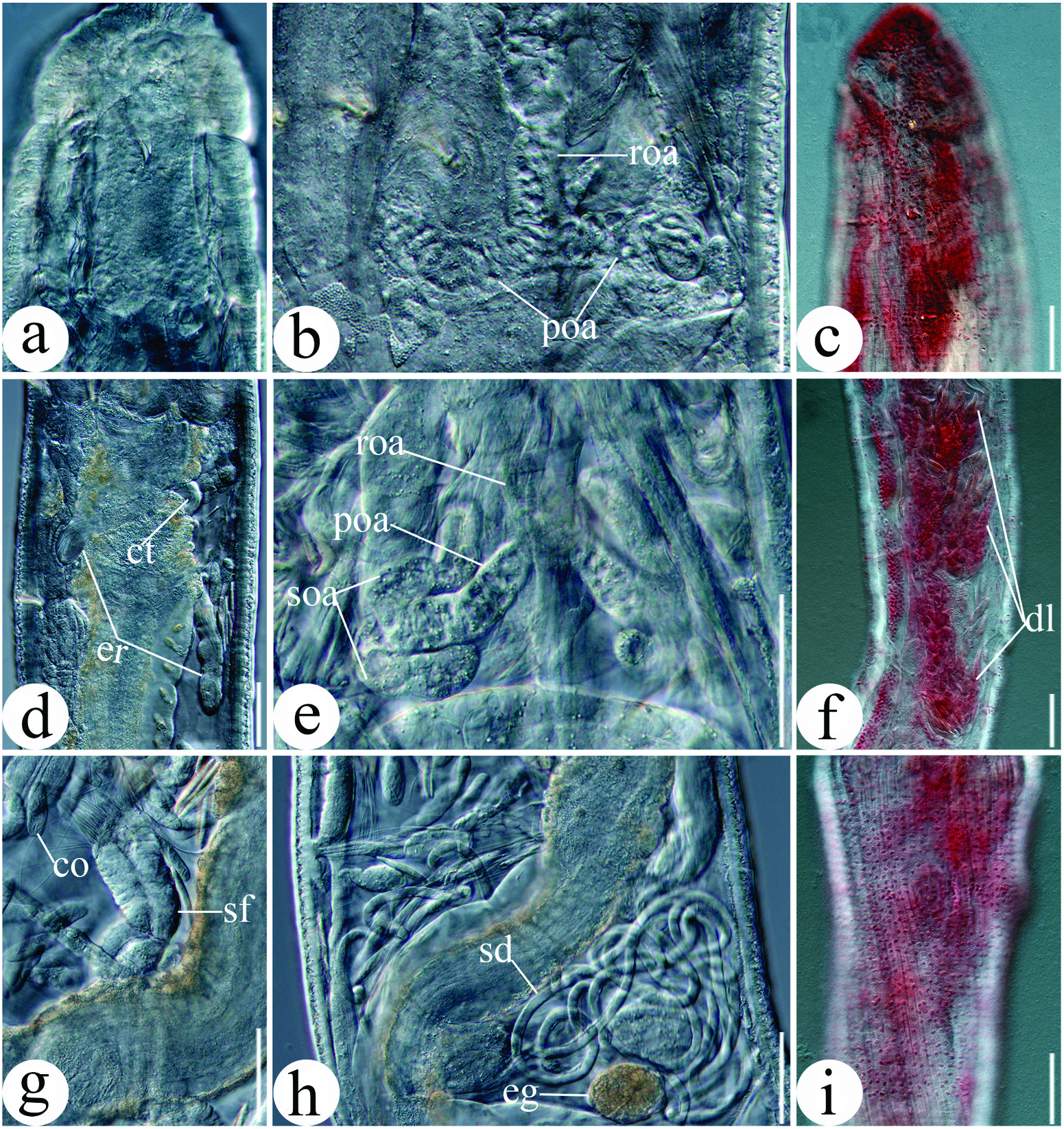
Small worms, transparent. Length 3.4–7.3 mm, diameter 0.12–0.21 mm at VII and 0.14– 0.23 mm at clitellum. Number of segments 30–43. Chaetae straight without inner hook, with slight proximal bend. Two chaetae per bundle throughout, absent in XII in mature worms ( Figure 6a View Figure 6 ). Ventral chaetae in anterior region ca. 25–37.5 μm long and 5 μm thick, chaetae in caudal region somewhat increasing in size, ca. 30–50 μm long and 5–6.5 μm thick. Lateral chaetae almost of equal size throughout, ca. 25–30 μm long and 5 μm thick. Head pore on prostomium mid-dorsally ( Figure 6a View Figure 6 ). Body wall 18–23 μm thick (in vivo), cuticle thin. Epidermal gland cells inconspicuous, 3–4 rows per segment, nearly rectangular, parallel or at chaetal level ( Figure 7c View Figure 7 ). Clitellum in XII–1/2XIII, scarcely thickening, girdle-shaped, hyalocytes and granulocytes in dense transverse rows ( Figures 6a View Figure 6 , 7i View Figure 7 ).
Brain ca 1.5× as long as wide (95 μm long, 65 μm wide, in vivo), deeply concave anteriorly, weakly incised posteriorly ( Figures 6a View Figure 6 , 7a View Figure 7 ). Oesophageal appendage with unpaired root dorsally behind pharyngeal pad in III, root with large lumen, extending posteriad, bifurcating into two primary branches, with meandering canal; each primary branch bifurcating further into two longer and thinner secondary branches, with smaller canal ( Figures 6a, b View Figure 6 , 7b, e View Figure 7 ). Three pairs of pharyngeal glands, poorly developed, pharyngeal glands in IV united dorsally, small, without ventral lobes, in V and VI dorsally separate, with ventral lobes ( Figures 6a View Figure 6 , 7f View Figure 7 ). No secondary pharyngeal gland lobes. Dorsal vessel beginning behind clitellum (segment XIV), blood colourless.
Three pairs of preclitellar nephridia from 6/7 to 8/9 ( Figure 6a View Figure 6 ), each about 115 μm long and 42 μm wide (in vivo). Anteseptale globular, consisting of funnel and parts of nephridial body, with coils of canal, postseptale elongate, length ratio of anteseptale: postseptale ca 1:1.3. Efferent duct originating from the middle of postseptale. Coelomocytes elongate, elliptical, numerous, 34–42 μm long, 15–18 μm wide (in vivo) ( Figure 7g View Figure 7 ).
Seminal vesicle absent. Sperm funnel cylindrical, tapering distally, ca. 106 μm long and 36 μm wide at collar (in vivo). Collar indistinct, as wide as funnel body, canal conspicuous ( Figures 6c View Figure 6 , 7g View Figure 7 ). Spermatozoa sparse, short, ca. 150 μm long, head ca. 18 μm long (in vivo) ( Figures 6c View Figure 6 , 7g View Figure 7 ). Diameter of sperm ducts ca. 6 μm, long and coiling unregularly in XII ( Figure 7h View Figure 7 ). Male copulatory organs muscular, male glandular body oval, ca. 55 μm long and 23 μm wide ( Figure 6a View Figure 6 ). Spermathecae free, not attached to oesophagus. Ectal pores lateral at 4/5, without ectal glands. Ectal ducts muscular, short, ca. 54 μm long and 10 μm wide (in vivo), with distinct ampullar dilatations in V (ca. 12 μm in diameter, in vivo); ampullar connecting tubes thinner than ectal ducts (ca. 8 μm wide, in vivo), extending into VII or VIII, ending in ellipsoid ental reservoirs, ca. 92 μm long and 19 μm wide (in vivo), ental reservoirs thin-walled, with spermatozoa ( Figures 6a View Figure 6 , 7d View Figure 7 ). One mature egg at a time.
Remarks
Among Hemienchutraeus species, there are six members with the same bifurcate branching pattern of the oesphageal appendage: H. bifurcatus Nielsen and Christensen, 1959 , H. csuzdii Dózsa-Farkas, 1989 , H. jeonjuensis Dózsa-Farkas and Hong, 2010 , H. planisetosus Xie et al., 1999 , H. solimoensis Righi, 1978 and H. tanjae Schmelz et al., 2005 . Among them, the new species is most similar to H. jeonjuensis in possessing three pairs of preclitellar nephridia from 6/7 to 8/9, spermathecae extending to VII–VIII, a girdle-shaped clitellum and the absence of a seminal vesicle. However, in H. jeonjuensis , there are two pairs of secondary pharyngeal glands in V and VI, respectively, and the dorsal vessel originates in preclitellar segments (X–XI).
The new species resembles H. csuzdii in the absence of secondary pharyngeal glands and a seminal vesicle, and in the postclitellar origination of the dorsal vessel. Conspicuous morphological differences from H. csuzdii include a short spermatheca confined to V and the first pair of preclitellar nephridia in 5/6. Hemienchytraeus planisetosus , H. solimoensis and H. tanjae differ from the new species in their longer spermathecae, presence of secondary pharyngeal glands, and the number and position of preclitellar nephridia.
The comparison of the new species with H. bifurcatus is difficult due to the lack of several key morphological traits in the original description of the latter; for example, nothing is known about the pharyngeal glands and the preclitellar nephridia. Schmelz et al. (2005) considered H. bifurcatus to be a species inquirenda since its type material is lost. Repeated attempts to collect specimens at the type locality have so far not been successful (Schmelz, unpublished data). However, it is unlikely that H. tenuiculus sp. nov. and H. bifurcatus are the same species: the segments are fewer in the latter, despite some overlap with the former (28–32), and its clitellum is said to be ‘strongly elevated’ ( Nielsen and Christensen 1959), while it is flat in the new species. Another difference can be inferred from the original illustration of H. bifurcatus ( Nielsen and Christensen 1959, fig. 27), which shows a brain with length:width ratio of 1:1. Furthermore, H. bifurcatus has been recorded only in Europe [ Denmark, Germany and Poland ( Schmelz and Collado 2010)].
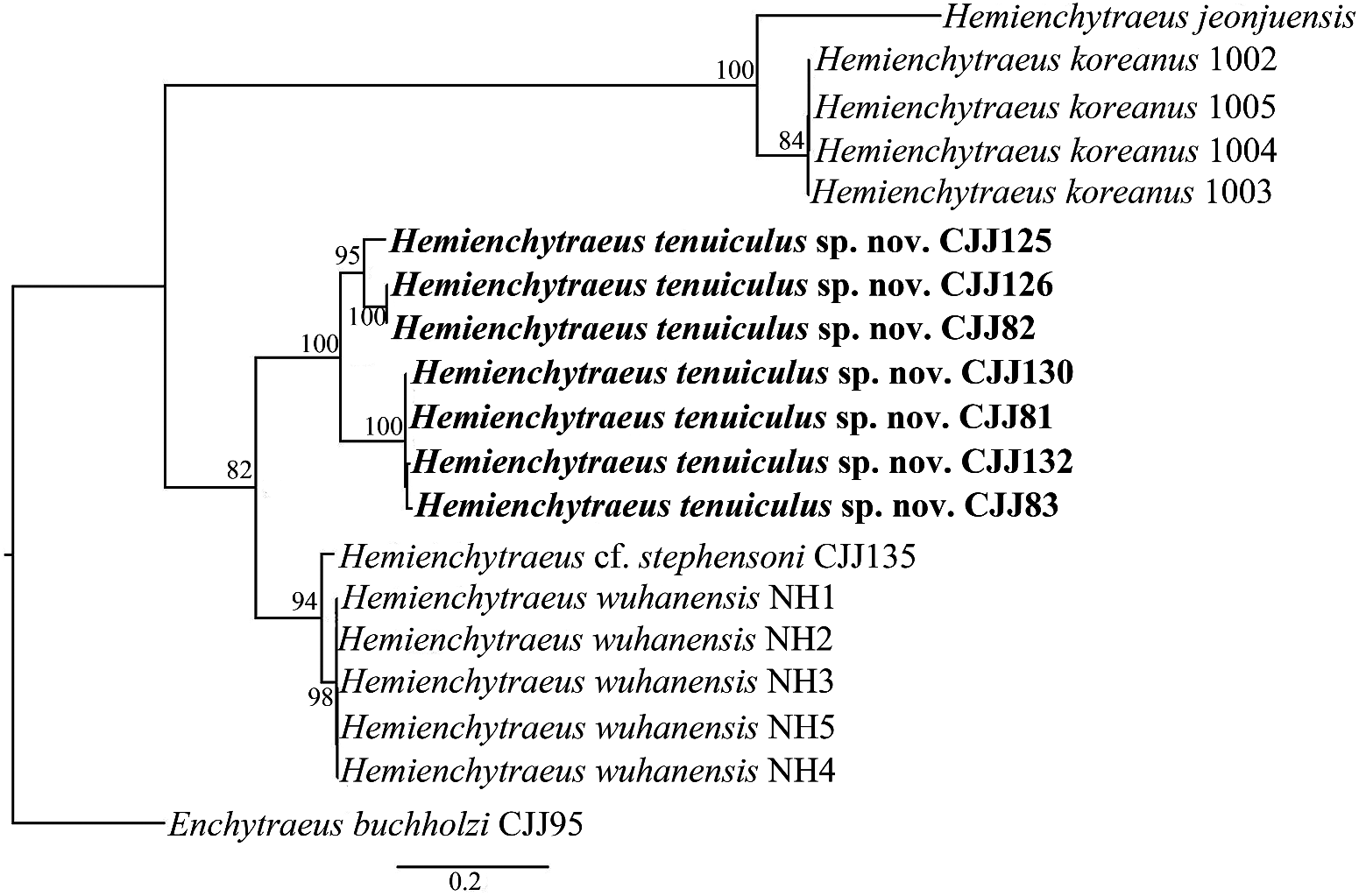
Hemienchytraeus tenuiculus sp. nov. is also clearly different from two species included in the DNA sequence comparison ( Figure 8 View Figure 8 ), H. koreanus Dózsa-Farkas and Hong, 2010 and H. wuhanensis Chen et al., 2021 . In both species each primary branch of the oesophageal appendage divides into more than two secondary branches. Furthermore, the two species have three pairs of secondary pharyngeal glands and five pairs of preclitellar nephridia, from 5/6 to 9/10. It is noteworthy that the morphological similarity of the new species and H. jeonjuensis is not reflected in the DNA sequences ( Figure 8 View Figure 8 ).
Phylogenetic analysis of the concatenated sequences (COI, H3 and ITS) shows that H. tenuiculus sp. nov. is monophyletic with respect to the other three species of Hemienchytraeus included in the analysis ( Figure 8 View Figure 8 ). However, the clade of the new species includes three branches, which suggests cryptic diversity within this morphospecies. For a detailed analysis see below (in the section on Molecular species delimitation).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |