
Sphaerodoridium guerritai, Moreira, Juan & Parapar, Julio, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3911.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:9B2EF89A-8915-4F9D-BBC3-EBE56E8011CE |
|
DOI |
https://doi.org/10.5281/zenodo.5671776 |
|
persistent identifier |
https://treatment.plazi.org/id/3C32A47A-FFEF-7329-FF3B-20816A22E3D1 |
|
treatment provided by |
Plazi |
|
scientific name |
Sphaerodoridium guerritai |
| status |
sp. nov. |
Sphaerodoridium guerritai View in CoL sp. nov.
( Figs. 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Material examined. Type material. Holotype: complete specimen, male, 5 mm long, 0.7 mm wide, with 27 chaetigers ( IINH 33095; BIOICE sample 2619; 67º16'86''N; 16º37'77''W; 600 m depth).
Allotype: complete specimen, female, 3.75 mm long, 0.75 mm wide, with 24 chaetigers ( IINH 33096; BIOICE sample 2666; 67º03'07''N; 16º16'90''W; 382 m depth).
Paratypes: BIOICE sample 2004 (28 spec., IINH 33097); 2046 (161, IINH 33098); 2360 (30, IINH 33099); 2480 (171, IINH 33100); 2526 (6, IINH 33101); 2606 (112, IINH 33102); 2619 (114, IINH 33103); 2622 (20, IINH 33104); 2652 (22, IINH 33105); 2666 (5, IINH 33106); 2019 (77, MNCN 16.01/16091).
Non-type material. Icelandic Museum of Natural History: BIOICE sample 2008 (1 spec.); 2009 (4); 2010 (18); 2011 (10); 2014 (1); 2024 (3); 2029 (4); 2030 (1); 2033 (6); 2040 (6); 2045 (88); 2047 (38); 2049 (4); 2053 (230); 2061 (38); 2062 (312); 2064 (35); 2067 (229); 2070 (36); 2087 (246); 2096 (16); 2097 (1); 2108 (2); 2113 (3); 2114 (18); 2117 (1); 2118 (11); 2119 (34); 2121 (4); 2124 (49); 2131 (35); 2132 (44); 2135 (1); 2136 (44); 2137 (22); 2139 (1); 2143 (5); 2145 (1); 2146 (6); 2149 (4); 2172 (23); 2304 (4); 2319 (39); 2325 (36); 2362 (55); 2364 (67); 2367 (1); 2481 (7); 2491 (1); 2504 (1); 2527 (43); 2539 (2); 2540 (114); 2542 (3); 2545 (3); 2548 (41); 2551 (1); 2554 (1); 2570 (2); 2572 (1); 2573 (21); 2575 (2); 2583 (1); 2585 (76); 2588 (2); 2589 (48); 2591 (5); 2594 (1); 2595 (1); 2613 (9); 2616 (19); 2627 (4); 2628 (40); 2629 (34); 2638 (1); 2655 (22); 2660 (19); 2669 (2); 2675 (1); 2682 (175); 2736 (2); 2741 (5); 2762 (1); 2783 (43); 2786 (1); 2787 (2); 2969 (1); 3033 (10); 3036 (35); 3099 (2); 3104 (5); 3108 (4); 3115 (2); 3247 (1); 3249 (1); 3252 (1); 3253 (22); 3660 (1); 3663 (53); 3669 (9).
Diagnosis. One transversal row of 11–12 stalked macrotubercles per chaetiger in midbody. Macrotubercle stalk about as long as or slightly longer than macrotubercle, with 1–3 small papillae along proximal half. Papillae on all body surfaces. 10–16 spherical papillae in front of each row of macrotubercles, somewhat arranged in a dorsal zig-zag row; 10–18 ventral papillae per chaetiger, most arranged in three more or less defined transversal rows: two on parapodial areas and one on interparapodial area, of usually 4, 6 and 5 papillae respectively. Females with a pair of oval, distally-opened tubercle-like structures located ventrally between parapodia of chaetigers 9–10. Males lacking any visible copulatory organs.
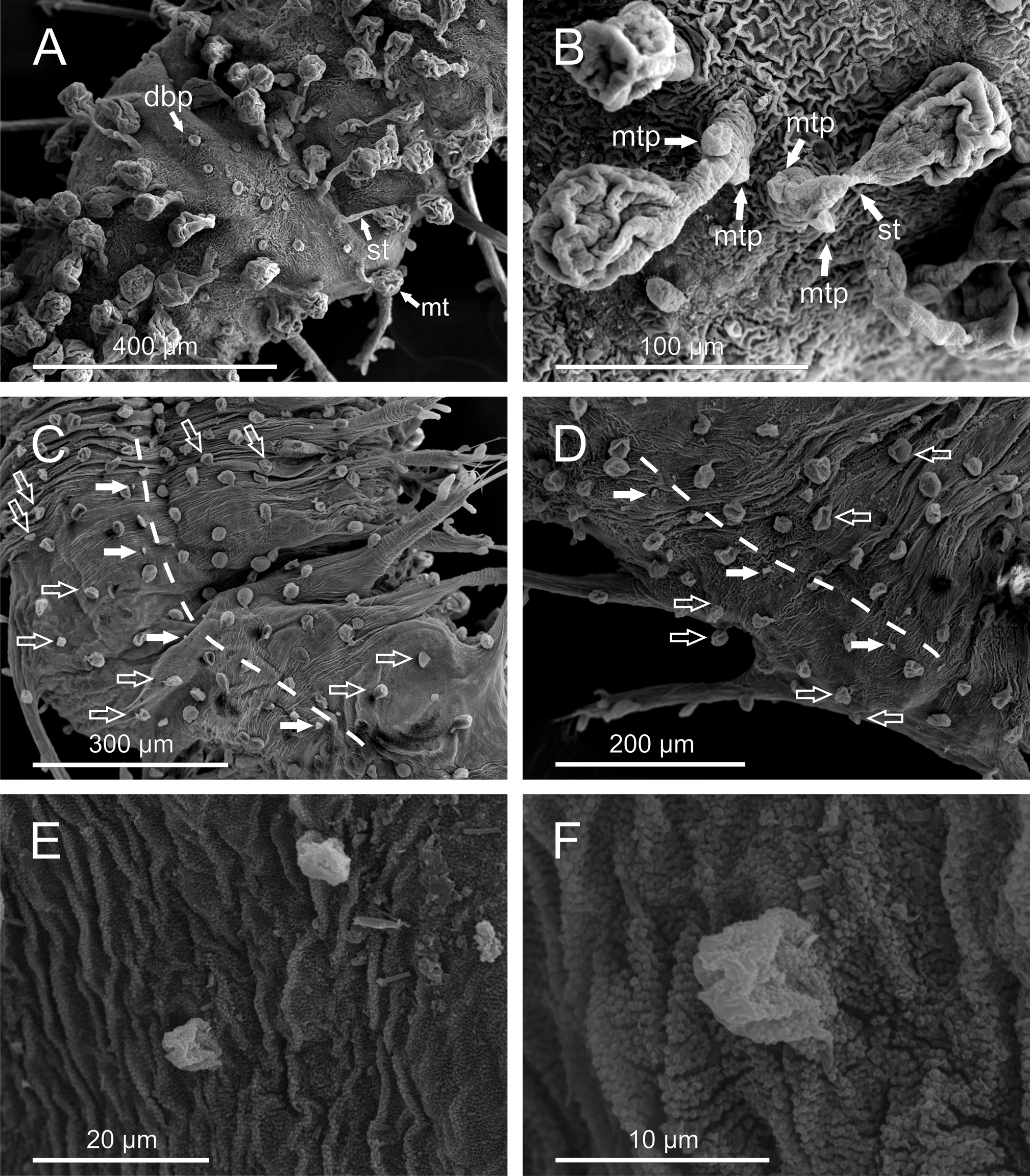
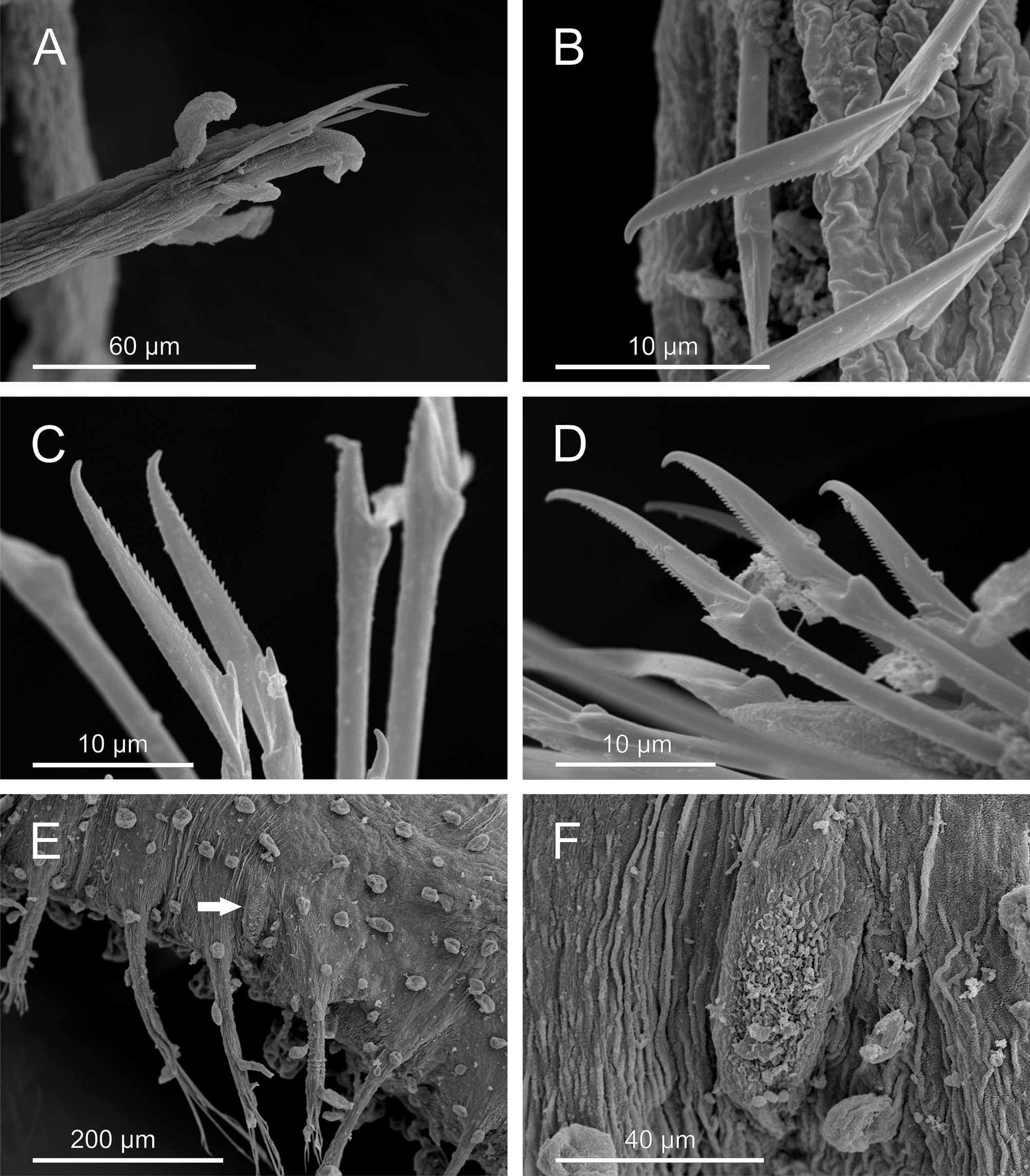
Description. Body of grub-like appearance, elongated ( Figs. 1 View FIGURE 1 A–B). Longest non-contracted preserved specimens measuring 5–7 mm in length, 0.37-0.70 mm in width (excluding parapodia), with 24–32 chaetigers. Longest contracted specimens measuring 5–6.4 mm in length, 0.75–1 mm in width, with 25–35 chaetigers. Preserved specimens lacking pigmentation, transparent-whitish. Tegument with a granulated appearance as observed under SEM ( Fig. 5 View FIGURE 5 F).
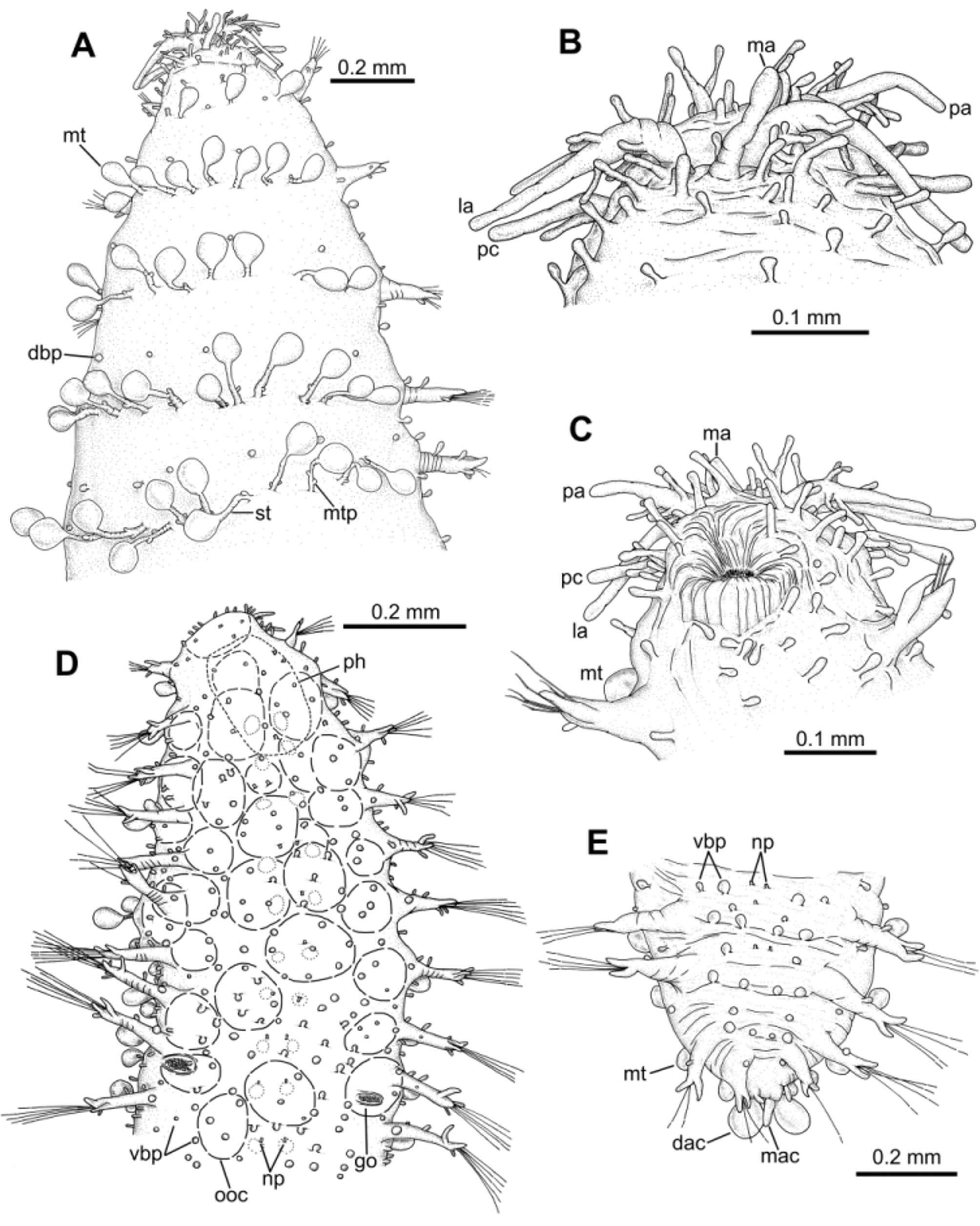
Prostomium and segments indistinctly separated from each other ( Fig. 2 View FIGURE 2 A). Prostomium bluntly rounded. Prostomial appendages: median antenna, one pair of lateral antennae and one pair of palps, all digitiform, elongated ( Figs. 2 View FIGURE 2 B–C). Median antenna smooth, about half of length of lateral antennae. Lateral antennae clearly longer than palps; palps inserted ventrally to lateral antennae. Lateral antennae and palps provided each with 7–10 (6–10) digitiform papillae (“spurs”) along proximal half. Prostomium and peristomium with numerous digitiform elongated papillae. One pair of peristomial cirri, digitiform, slightly shorter than palps, with two elongated papillae on proximal third, one dorsal and one ventral; six papillae surrounding mouth opening ( Fig. 2 View FIGURE 2 C). Two dark eyes; visible dorsally from prostomium to chaetiger 2. Pharynx extending over 3 chaetigers (3–4).
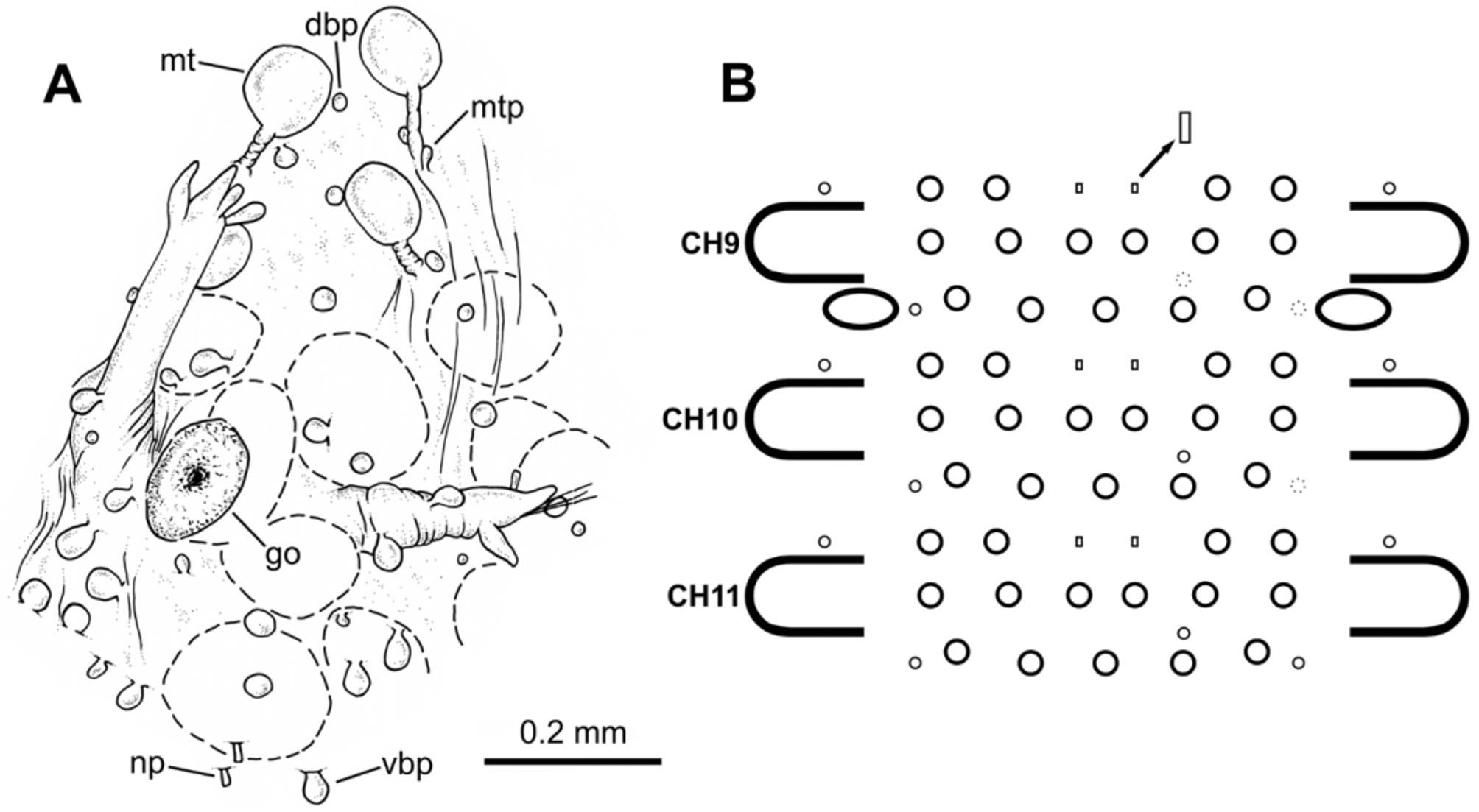
Dorsal macrotubercles stalked, slightly spherical to oval ( Fig. 5 View FIGURE 5 B); stalk long, about as long as or slightly longer than macrotubercle; all stalks provided with 1–3 (usually two) small oval to spherical papillae along proximal half; stalk papillae smaller than body papillae. Contracted specimens with macrotubercle stalk appearing shorter, about 1/2–2/3 length of macrotubercle. Macrotubercles arranged in one transversal dorsal row per chaetiger: 5 macrotubercles on chaetiger 1 (4–6), 8 on chaetigers 2–3 (8–11), 11–12 on following chaetigers and 6 on last chaetiger. About 10–16 spherical papillae in front of each macrotubercle row, somewhat arranged in a dorsal zig-zag row ( Fig. 1 View FIGURE 1 C); 2–4 papillae on lateral chaetiger surfaces (i.e. surfaces between insertion of parapodia and lateralmost macrotubercle), one of them located in between two consecutive parapodia. Ventral surface of each chaetiger with 10–18 papillae of different sizes, usually with short peduncle (<1/3 length of papilla); shape of papillae (excluding peduncle) elongated to spherical; most of them arranged following a non-random pattern, more or less defining three transversal rows: two on parapodial areas and one on interparapodial area, of usually 4, 6 and 5 papillae respectively; other smaller papillae sometimes present at both sides of third row (interparapodial) and among second and third row ( Figs. 4 View FIGURE 4 B, 5C). Each chaetiger (except first and last 2–3) with one pair of short tubular papillae, each with distal opening, interpreted as nephridial papillae and nephridiopore, respectively ( Figs. 5 View FIGURE 5 D–F); nephridial papillae located on ventral surfaces at midlength of a row of four body papillae, slightly in front of parapodial areas.
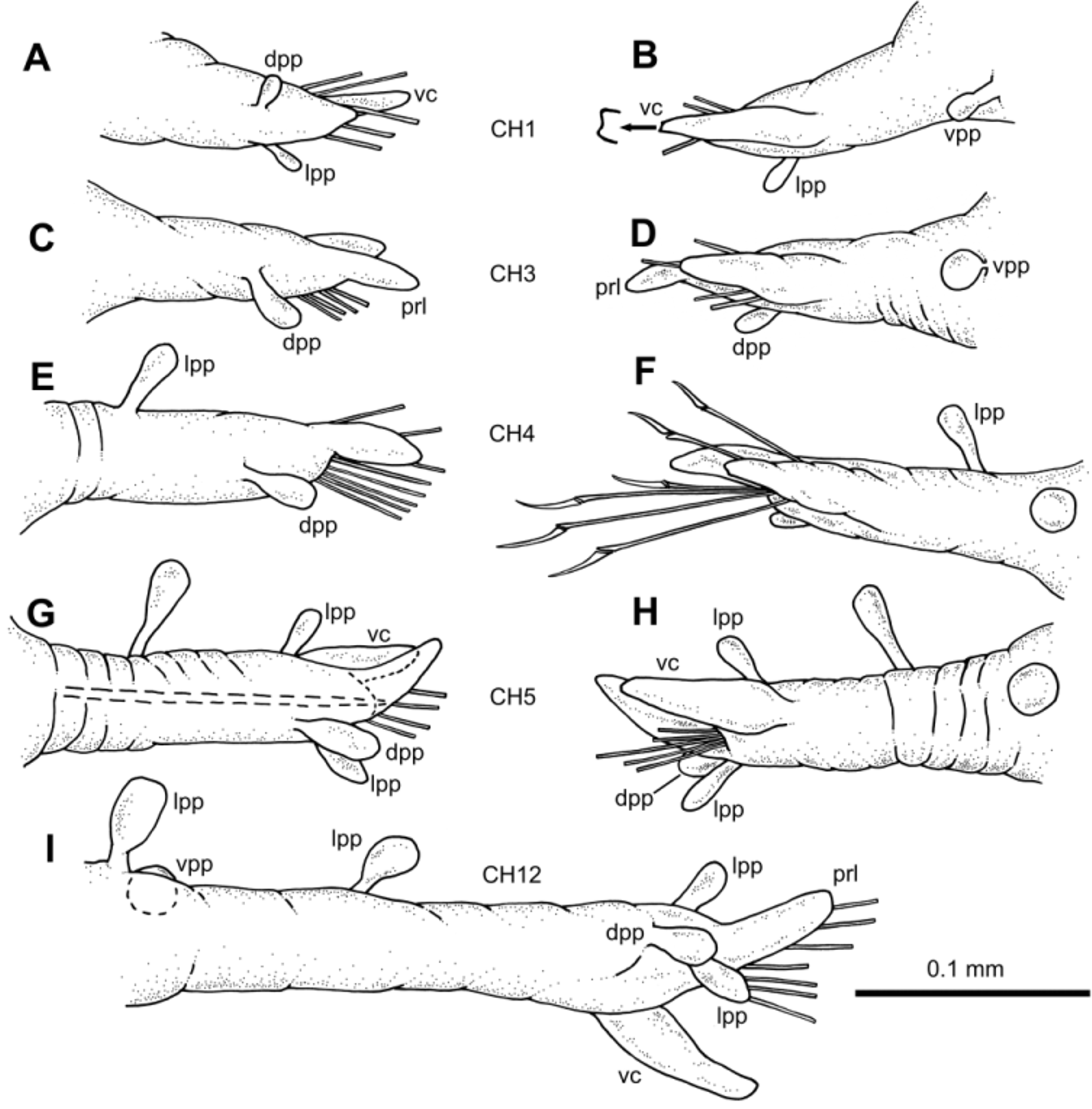
Parapodia unirramous, usually with wrinkled surface, much longer than wide ( Figs. 3 View FIGURE 3 A–I, 6A). Chaetiger 1–2 parapodia shorter than following, lacking prechaetal lobe. Following parapodia with digitiform prechaetal lobe, projecting well beyond acicular lobe. Ventral cirri digitiform, inserted at distal third, surpassing acicular lobe, shorter than prechaetal lobe; distal end appearing slightly bifid at higher magnification. Parapodia increasing in length steadily from about chaetiger 10 backwards; last chaetiger parapodia shorter than preceding ones. Parapodial papillae elongated, with basal peduncle; shape (excluding peduncle) spherical to oval. Midbody parapodia with 5–6 papillae along parapodial lobe; distal third with up to three papillae: one on anterior lateral surface, one on dorsal surface and one on posterior lateral surface; two papillae on lateral anterior surface of parapodium proximal half; one papilla with shorter peduncle on ventral proximal surface, more spherical in shape than others. Chaetigers 1–4 with 2–3 papillae.
One thick acicula per parapodium, distal end pointed. Compound falciger chaetae numbering 4–15 per fascicle ( Figs. 6 View FIGURE 6 B–D); shaft distal end inflated with thin spinulation; blades unidentate with recurved tip and thin spinulation along most of cutting edge. Blades showing gradation in length within same fascicle (22–32 µm anterior parapodia; 25–27 µm midbody parapodia; 20–25 µm posterior parapodia).
Pygidium terminal, with midventral digitiform anal cirrus, flanked dorsally by two shorter elongated papillae; one pair of dorsal anal cirri similar to dorsal macrotubercles of posterior chaetigers ( Figs. 1 View FIGURE 1 D, 2E).
Holotype male carrying spermatozoa, visible through body wall along most of body; spermatozoa conspicuously stained when treated with methylene blue; holotype and paratype males lacking ventral special copulatory structures. Females carrying visible oocytes; with a pair of tubercle-like, distally opened structures, interpreted as genital openings, located ventrally between parapodia of chaetigers 9–10 ( Figs. 2 View FIGURE 2 D, 4A, 6E–F). Sex ratio roughly 1:1 (n = 607; 30 samples).
Etymology. The new species is named after our colleague and friend, Dr. José M. Guerra-García ("Guerrita") from Universidad de Sevilla ( Spain), in regard of his many contributions to the field of Marine Biology and his friendship.
Ecology and distribution. Most of the specimens were found in waters around the northern half of Iceland, between parallels 64º and 68º (apart from those of sample 2304 -SW Iceland- and 2325 -SE coast-), at depths of between 49–1253 metres, mostly in muddy sediments (sandy silt and silt), with a range of temperature from -0.86 to 8.04ºC. Samples with the largest numbers of specimens (100–312) were collected with the RP sledge, at depths of 82– 735 m.
Remarks. The new species has been assigned to the genus Sphaerodoridium because of the presence of stalked macrotubercles and the short length of the median antenna when compared to that of the lateral antennae ( Fauchald 1974). Sphaerodoridium guerritai sp. nov. mostly differs, however, from all species in the genus by having dorsal macrotubercles which are provided with a long stalk which bears, in turn, 1–3 small papillae along its proximal half. In the NE Atlantic, two species of Sphaerodoridium have been reported, namely S. claparedii ( Greeff, 1866) and S. fauchaldi Hartmann-Schröder, 1993 . Apart from the aforementioned features of the macrotubercle stalk, those species differ from S. guerritai sp. nov. in having up to 6 macrotubercles per dorsal row in midbody instead of 11–12 and less parapodial papillae (1–3 vs. 5–6); furthermore, S. fauchaldi lacks papillae on dorsal body surfaces ( Table 1 View TABLE 1 ).
Three further species of Sphaerodoridium have been reported from other parts of the world: S. japonicum Ozolin’sh, 1987 (Northwest Pacific), S. lutzeni Kudenov, 1987 (Gulf of Mexico) and S. campanulata Borowski, 1994 (Southeast Pacific). Of these, S. lutzeni and S. japonicum bear less macrotubercles per dorsal row than S. guerritai sp. nov. (6–8 vs. 11–12), S. japonicum and S. lutzeni lack dorsal body papillae, and S. campanulata has conspicuous dorsal bell-shaped papillae similar in size to the dorsal macrotubercles. In addition, Sphaerodoridium species seem to differ in the presence and number of papillae on the prostomial appendages. For instance, S. japonicum and S. campanulata have lateral antennae and palps which lack papillae (Ozolin’sh 1987; Borowski 1994; Böggemann 2009) while the remaining species show 1–2 papillae per appendage ( S. claparedii , S. fauchaldi , S. lutzeni ) or 6–10 ( S. guerritai sp. nov.). Peristomial cirri are instead usually smooth ( S. claparedii , S. japonicum , S. lutzeni , S. fauchaldi , S. campanulata ) whereas in S. guerritai sp. nov. each cirrus bears two papillae. The number and size of ventral body papillae also show different patterns across the genus: S. lutzeni and S. campanulata present fewer papillae (4–6) whereas the others show a greater number (10–18); ventral papillae are of different sizes in S. claparedii ( Southern 1914) and S. guerritai sp. nov., and of similar size in S. lutzeni , S. fauchaldi and S. campanulata .

| MNCN |
Museo Nacional de Ciencias Naturales |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.