
Hediste japonica, (IZUKA, 1908)
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2003.00059.x |
|
persistent identifier |
https://treatment.plazi.org/id/534787FA-FFBE-FFE6-9B4D-9674FA32A1F6 |
|
treatment provided by |
Carolina |
|
scientific name |
Hediste japonica |
| status |
|
HEDISTE JAPONICA ( IZUKA, 1908) View in CoL ( FIGS 2-15 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 View Figure 10 View Figure 11 View Figure 12 View Figure 13 View Figure 14 View Figure 15 , 38-41 View Figure 38 View Figure 39 View Figure 40 View Figure 41 , 43 View Figure 43 )
Nereis japonica Izuka, 1908: 295–305 View in CoL , 4 text figs.; Izuka, 1912: 163–169, pl. 17, figs 14-16, 18 and 4 text figs.
? Nereis japonica View in CoL (populations of Yellow Sea): Khlebovich et al., 1982: 1110–1113.
Neanthes japonica View in CoL (epitokes): Smith, 1958: 60–73.
Hediste sp. 2 (Ariake form): Sato, 1999: 129–143; Sato, 2000: 187–191, figs 8-1 and 8-3; Sato, 2001: 66–86.
Not Hediste japonica: Imajima, 1972: 102–105 View in CoL , figs 30 a- n, 37 (= H. diadroma View in CoL sp. nov. or H. atoka View in CoL sp. nov.); Imajima, 1996: 139, fig. 111 (= H. diadroma View in CoL sp. nov. or H. atoka View in CoL sp. nov.); Sato, 1999: 129–143 (= H. diadroma View in CoL sp. nov.); Sato, 2000: 187–191, figs 8-2 and 8-3 (= H. diadroma View in CoL sp. nov.); Sato, 2001: 66–86, figs 4-1-4-10 (= H. diadroma View in CoL sp. nov.); Rouse & Pleijel, 2001: pl. 5c (= H. diadroma View in CoL sp. nov.).
Type material ( Fig. 2 View Figure 2 )
Lectotype (UMUTZ-PLC-IZ-55A): complete ovigerous female ( BL: 100 mm, BW: 4.0 mm, 98 setigers), Hachihama, Kojima Bay, Okayama Prefecture, the Seto Inland Sea , 17 December 1906, coll. A. Izuka. A part of paralectotype (UMUTZ-PLC-IZ-55B): 7 ovigerous females ( BL: 70–100 mm, BW: 3.0– 4.1 mm, 87–102 setigers), data as for lectotype.
Other material examined
Mature specimens (epitokes): collected during reproductive swarming in Ariake Sea, Japan. Nanaura, Kashima-shi , Saga Prefecture, 29 December 2001, coll. A. Nakashima, 6 specimens (OMNH-Iv-4223, 4224) .
Epitokes obtained after rearing in laboratory: Daijugarami , Higashiyoka-cho, Saga Prefecture, Ariake Sea, Japan, 1 female fixed 24 February 1997 after rearing for 4 months (ZIHU-2102), 1 male fixed 20 January 1997 after rearing for 3 months (ZIHU-2103), coll. A. Nakashima.
Specimens collected from sediment samples: Seto Inland Sea, Japan: Kojima Bay , Okayama Prefecture, 13 October 1908, coll. Eitaro Iwaki, 7 specimens (damaged by drying, UMUTZ-PLC-IZ-0000) . Ariake Sea, Japan: Around mouth of Honmyogawa River , Isahaya Bay , Takaki-cho , Nagasaki Prefecture, 21 August 1994, coll. H. Yamashita, 5 specimens ; 19 November 1994, coll. H. Yamashita, 11 specimens ; 28 April 1995, coll. T. Sakai, 17 specimens (juveniles) ; 29 May 1995, coll. T. Sakai, 17 specimens ; 21 January 1996, coll. T. Sakai, 2 specimens (juveniles) ; 28 November 1996, coll. A. Nakashima, 31 specimens. Onojima, Isahaya Bay, Isahaya-shi , Nagasaki Prefecture, 30 March 1994, coll. M. Sato, 52 specimens (juveniles) ; 1 July 1994, coll. H. Yamashita, 13 specimens ; 20 September 1994, coll. H. Yamashita, 9 specimens; 28 October 1995, coll. T. Sakai, 1 specimen ; 10 November 1997, coll. M. Sato, 1 specimen. Around mouth of Nitandagawa River, Isahaya Bay , Moriyama-cho , Nagasaki Prefecture, 20 August 1994, coll. M. Sato, 12 specimens . Ariakegawa River, Isahaya Bay, Moriyama-cho , Nagasaki Prefecture, 20 August 1994, coll. M. Sato, 36 specimens . Yamadagawa River, Isahaya Bay, Azuma-cho , Nagasaki Prefecture, 24 April 1995, coll. H. Yamashita, 22 specimens (juveniles) . Kojirogawa River, Isahaya Bay , Nagasaki Prefecture, 3 November 1994, coll. H. Yamashita, 9 specimens . Nanaura, Kashima-shi , Saga Prefecture, 27 September 1996, coll. H. Yamashita, 1 specimen ; 29 April 2001, coll. M. Sato, 11 specimens (ZIHU-2104) . Rokkakugawa River, Ashikari-cho , Saga Prefecture, 10 October 1995, coll. H. Yamashita & T. Sakai, 11 specimens . Daijugarami, Higashiyoka-cho , Saga Prefecture, 20 March 1995, coll. A. Nakashima, 12 specimens (juveniles) ; 3 April 1995, coll. A. Nakashima, 5 specimens (juveniles) ; 28 July 1995, coll. A. Nakashima, 90 specimens (ZIHU-2101); 21 October 1996, coll. A. Nakashima, 21 specimens (CMNH-ZW-1888, MNHN, NSMT-Pol-107441, SMF-12064, USNM- 1008435, ZMUC-Pol-1502) ; 8 December 1996, coll. A. Nakashima, 16 specimens. Omutagawa River, Omutashi , Fukuoka Prefecture, 24 November 1998, coll. K. Ichimiya, 21 specimens ; 4 January 1999, coll. K. Ichimiya, 21 specimens (including mature or nearly mature adults) ; 17 May 1999, coll. M. Sato, S. Iwamatsu & A. Kubo, 32 specimens; 21 December 1999, coll. S. Iwamatsu, 5 specimens (including mature or nearly mature adults). Yellow Sea, Korea: Yongyudo Island , Inchon, 19 January 1999, coll. M. Sato & J.-S. Hong, 11 specimens (including nearly mature adults) . Around mouth of Kumu River, Kunsan , 2, 3 May 2000, coll. A. Kubo, 17 specimens .
Diagnosis
Few (less than 10 in most cases) large paragnaths on right and left sides of proboscis in group II. Homogomph falcigers with slender blade present, and heterogomph spinigers absent in neuropodia. Neuropodial postchaetal ligule tapering to digitate lobe throughout. Epitokous stage without any addition of epitokespecific chaetae. Full-grown oocytes 180–210 Mm in diameter. Reproductive swarming in winter.
Description
Body stout anteriorly, posteriorly gradually tapering toward pygidium ( Figs 3a,c View Figure 3 , 4a,d View Figure 4 ). Dorsum convex, venter relatively flat with longitudinal midventral groove. Colour in preserved specimens whitish cream with dark brown pigmentation on the anterior dorsal surface; colour in life at sexually immature stage reddish or greenish brown with anterior dorsal surface darkened.
Prostomium pyriform, broader than long, with pair of smooth tapered antennae situated at anterior end. Pair of palps with massive palpophores about twice as long as antennae and short round palpostyles. Two pairs of round or reniform eyes almost equal in size (border of eye unclear in type material because of discoloration from long-term preservation), arranged trapezoidally (space between anterior pair 1.3–1.4 times as wide as that between posterior pair). Longitudinal mid-dorsal groove present on anterior dorsum of prostomium. Partial dark pigmentation present on dorsal anterior surface of prostomium and palpophore ( Fig. 4a View Figure 4 ), though pigmentation unclear in type material ( Fig. 3a View Figure 3 ).
Peristomium nearly as long as following setigers, with 4 pairs of tentacular cirri of unequal length; posterior dorsal tentacular cirri longest, reaching back to setigers 6–9; anterior dorsal tentacular cirri next longest, reaching back to setigers 3–4.
Proboscis with pair of dark brown jaws, each with 8–9 teeth. Black paragnaths, usually with sharply pointed tip, present on both maxillary and oral rings; paragnath numbers in lectotype (range for all 10 mature specimens in parentheses) as follows. Group I: 2 (1–4); II: 6 on right and 11 on left side in two arched rows, total 17 (10–22); III: 28 (23–30) in transverse band; IV: 10 on right and 11 on left in two arched rows, total 21 (10–21); V: none; VI: 5 on right and 8 on left, in small clusters on gentle mound, total 13 (10–18); VII- VIII: 22 (19–26) in single transverse row. Paragnaths in group II conspicuously larger than the others, with hook-shaped tip. Two gentle mounds and 5 more proximal (on everted proboscis) conical mounds, resembling sometimes papillae, present as transverse rows parallel to paragnath row in group VII- VIII ( Fig. 4c View Figure 4 ).
Parapodia of first 2 setigers uniramous, all following parapodia biramous ( Figs 3d- j View Figure 3 , 4e- k View Figure 4 ). Uniramous parapodia with reduced notopodia consisting of dorsal cirrus and superior ligule, with ordinary neuropodia.
Notopodia consisting of dorsal cirrus and three ligules in biramous parapodia, i.e. large superior ligule and upper and lower acicular ligules; all notopodial ligules subtriangular with tapering tip. Upper acicular ligule subequal to lower one in anterior setigers ( Figs 3f View Figure 3 , 4g View Figure 4 ); gradually diminishing in size in middle and posterior setigers ( Figs 3g- j View Figure 3 , 4h- k View Figure 4 ). Superior ligule thick in anterior setigers, thinner in middle and posterior setigers, most expanded in middle setigers. Dorsal cirri slender, tapering, gradually diminishing in size in posterior setigers.
Neuropodia consisting of ventral cirrus and three ligules throughout, i.e. prechaetal acicular ligule, postchaetal ligule and inferior ligule. Prechaetal acicular ligule and postchaetal ligule conical, of similar length, completely separate in anterior setigers (up to about first 10 setigers), fused at basal parts with their tapering tips remaining in middle and posterior setigers ( Figs 3h,j View Figure 3 , 4i,k View Figure 4 ). Inferior ligule conical. Ventral cirrus slender with tapering tip. Inferior ligule and ventral cirrus largest in anterior setigers, gradually diminishing in size in middle and posterior setigers.
Notochaetae all homogomph spinigers ( Figs 5a View Figure 5 , 6a View Figure 6 , 7a,b View Figure 7 ); terminal sections (blades) very slender with finely serrated cutting edge. Numbers of notochaetae of atokes 10–40 in anterior and middle setigers, and less than 10 in posterior setigers in most specimens.
Neurochaetae in supra-acicular fascicle consisting of homogomph spinigers (around 10 in most cases, Fig. 5b View Figure 5 ) and heterogomph falcigers (up to 10 in most cases, Figs 5b View Figure 5 , 6b View Figure 6 , 7c View Figure 7 ) in anterior setigers. Neuropodial homogomph spinigers similar to notopodial ones in shape and size. Heterogomph falciger relatively thick, with serrated blade present in anterior and middle setigers (anterior 50–60 setigers in adults), and 1–3 thick simple chaetae with tapering tip ( Figs 6c View Figure 6 , 7d View Figure 7 ) present instead of heterogomph falcigers in posterior setigers ( Fig. 5b View Figure 5 ). Incomplete simple chaetae found in several setigers (around setiger 40) just prior to posterior setigers with complete simple chaetae in juvenile, indicating formation of simple chaeta by deformation of tip of stalk of heterogomph falciger after detachment of its blade ( Fig. 8 View Figure 8 ).
Neurochaetae in infra-acicular fascicle consisting of homogomph spinigers (5–20 in most cases, Figs 5c View Figure 5 , 6d View Figure 6 ) at upper position, homogomph falcigers (up to 20 in most cases, Figs 6e View Figure 6 , 7e View Figure 7 ) on middle portion, and heterogomph falcigers (up to 10 in most cases, Fig. 6f View Figure 6 ) at lower position; homogomph and heterogomph falcigers relatively thin with a slender finely serrated blade, often indistinguishable between homogomph and heterogomph.
Acicula black except colourless proximal part; single acicula present in each ramus (occasionally 2 or 3 aciculae present in each ramus).
Pygidium with anus on dorsal side, with pair of cylindrical slender anal cirri ( Figs 3c View Figure 3 , 4d View Figure 4 ).
Epitokous metamorphosis at sexually mature stage: body colour changed to green in both males and females. Eyes slightly enlarged ( Fig. 9a,b View Figure 9 ). Parapodial ligules (especially upper and lower acicular ligules of notopodia) enlarged slightly in both males and females ( Fig. 9c,d View Figure 9 ); dorsal and ventral cirri also markedly elongated in males. Homogomph spinigers increasing in number (up to about 100 in notopodial fascicles, and up to about 50 each in neuropodial supra-acicular and infra-acicular fascicles) in some specimens ( Figs 6g View Figure 6 , 7a,b View Figure 7 ).
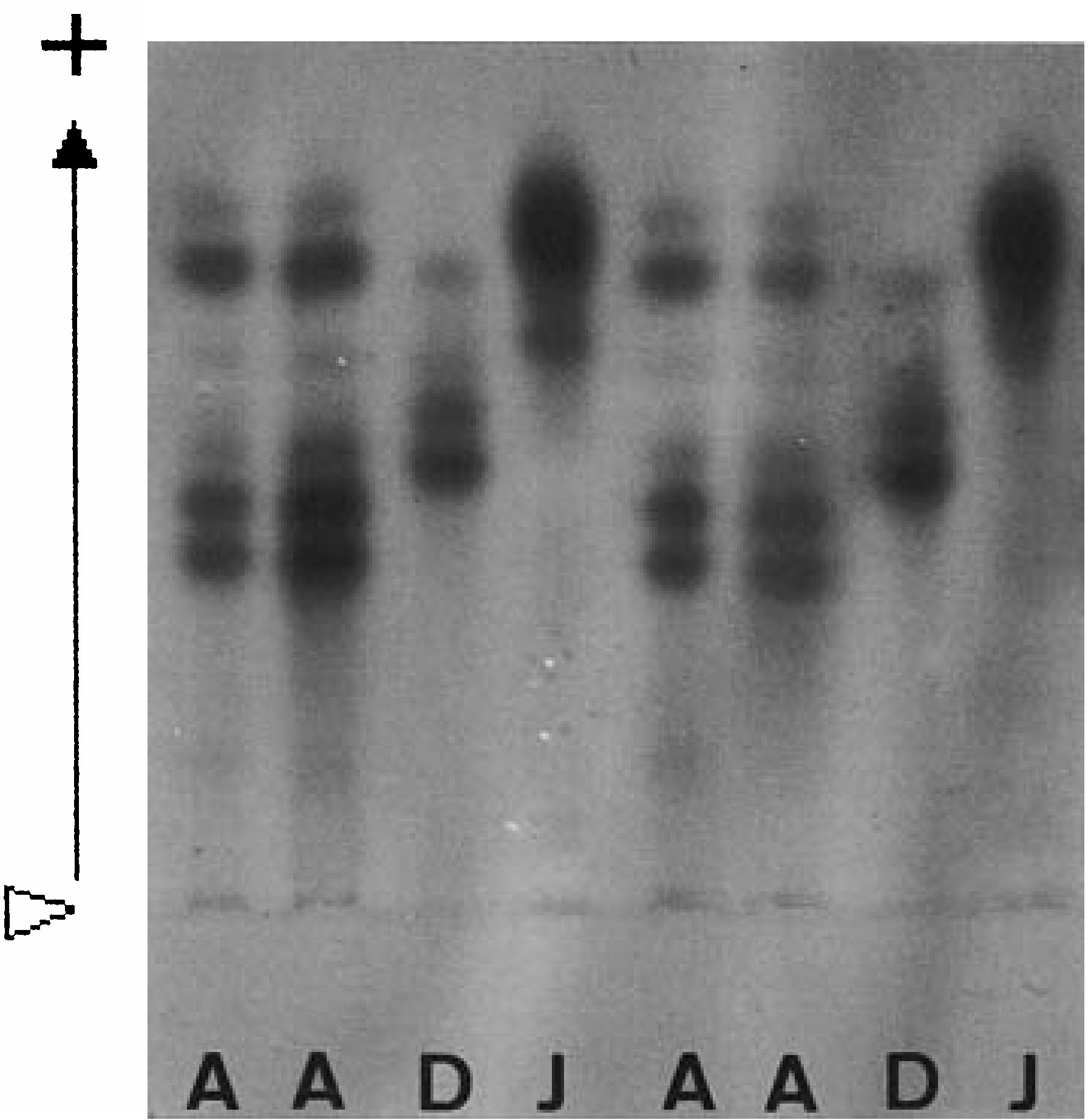
Allozyme pattern of LDH
Two bands of anodal migration, the wider more mobile and the narrower less mobile, were observed in all specimens examined (31 from Isahaya Bay, 88 from Daijugarami, 42 from Omutagawa River) ( Fig. 10 View Figure 10 ). No variation of band pattern was observed, indicating that the loci for the allozymes of LDH were monomorphic.
Allometry
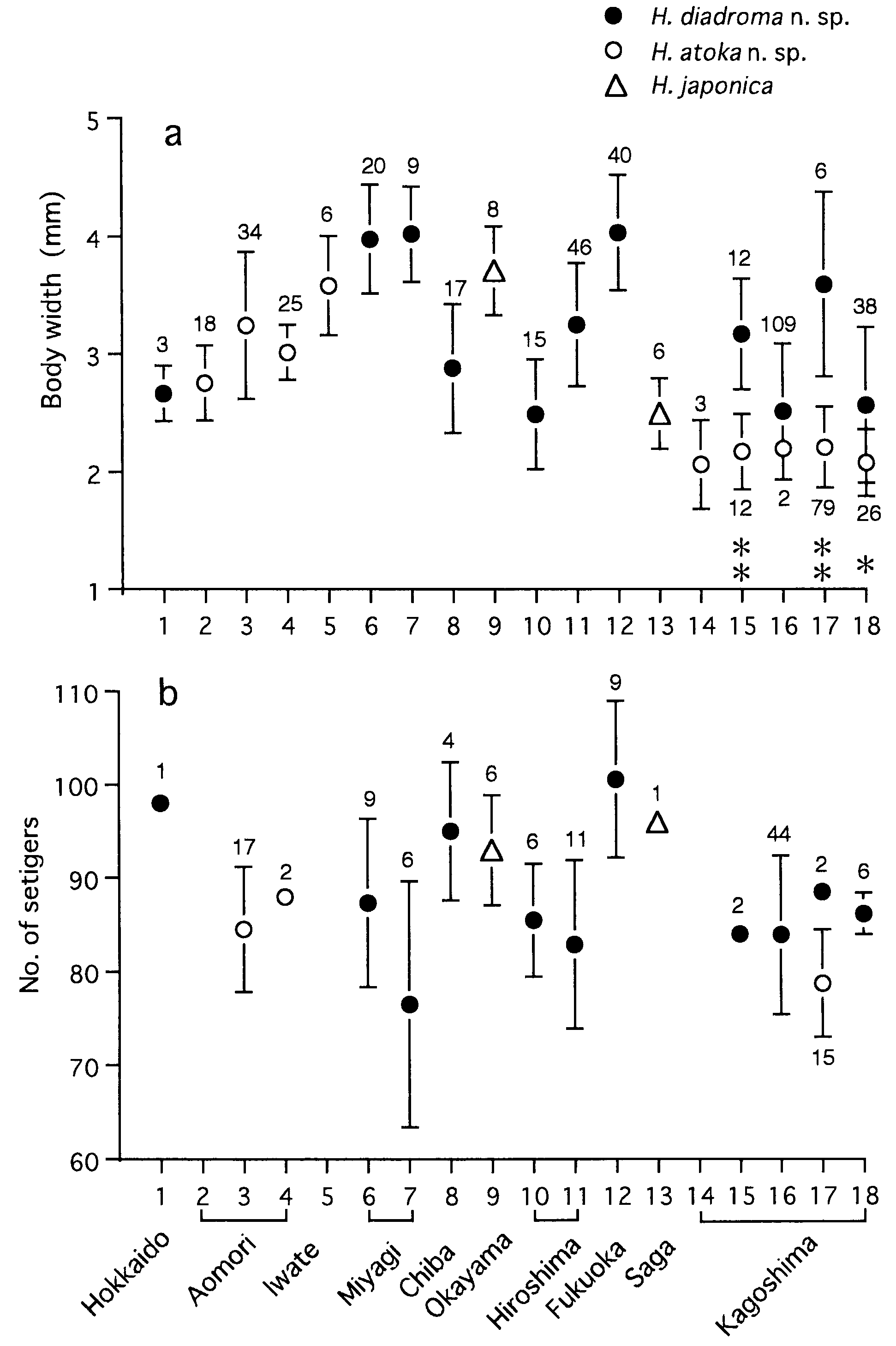
Body length ( BL mm) was well correlated with body width ( BW mm) according to the regression formula ( Fig. 11a View Figure 11 ): BL = 29.4 BW - 17.9 (r 2 = 0.95, P <0.0001, n = 51). Setiger number (SN) was correlated with BW, according to the regression formula with a high gradient in smaller specimens, i.e. juveniles (less than 2 mm in BW) ( Fig. 11b View Figure 11 ): SN = 53.3 BW – 16.3 (r 2 = 0.64, P <0.0001, n = 22). However, in the larger specimens, i.e. adults (2 mm or more in BW), setiger number was rather constant at around 90–100, according to the regression formula with a low gradient: SN = 4.5 BW + 78.4 (r 2 = 0.17, P = 0.03, n = 27). Maximum body sizes were 125 mm in BL, 4.9 mm in BW, and 108 in SN.
Eye width ( EW mm) was correlated with BW according to the regression formula in immature specimens: EW = 0.03 BW + 0.04 (r 2 = 0.66, P <0.0001, n = 46) ( Fig. 12 View Figure 12 ). EW (0.16–0.19 mm) of mature adults (n = 8) was significantly larger than EW (0.10–0.16) of immature specimens (n = 7) with corresponding BW of 2.1– 2.9 mm (Mann–Whitney U -test, P = 0.002).
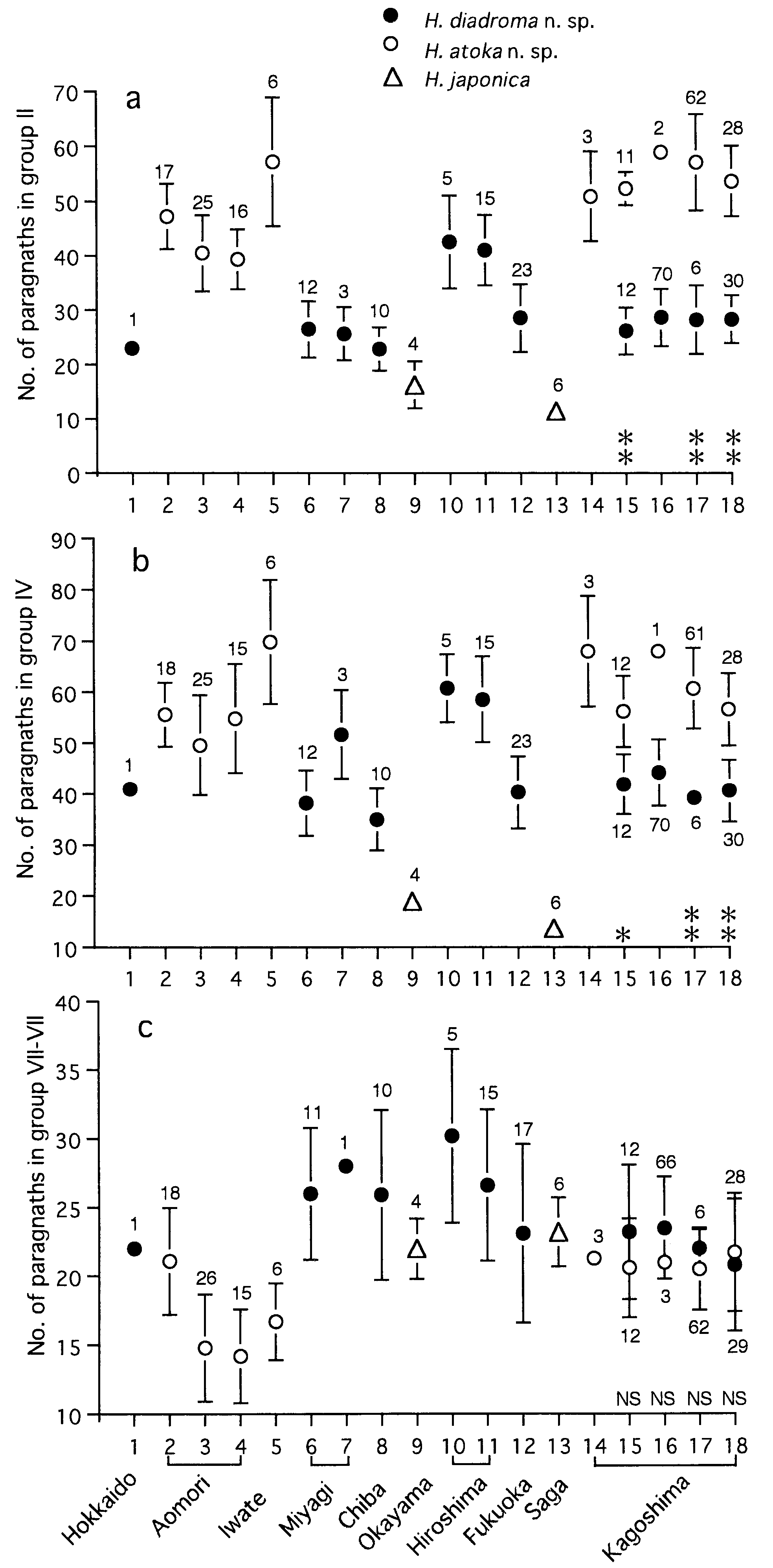
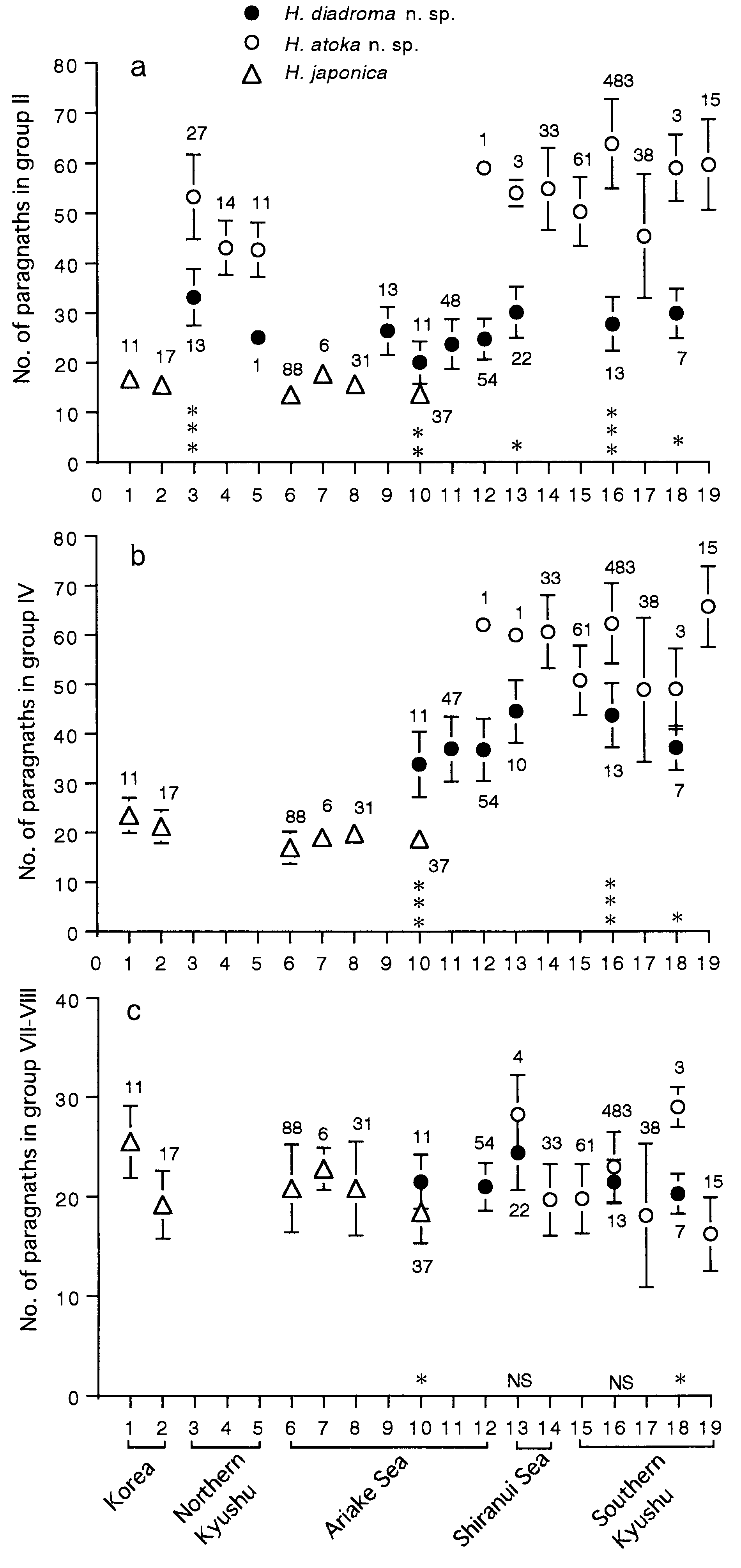
Paragnath number ( PN) in each group increased rapidly with correlation to increasing body width in juveniles, with a high correlation coefficient and a high gradient of its formula, e.g. PN = 8.3 BW + 0.5 (r 2 = 0.53, P <0.0001, n = 22) for group II, PN = 26.2 BW - 14.4 (r 2 = 0.58, P <0.0001, n = 21) for group III, PN = 74.3 BW - 30.3 (r 2 = 0.58, P <0.0001, n = 21) for all groups, but rather stable in adults, with a low correlation coefficient and a low gradient of its regression formula, e.g. PN = 1.0 BW + 11.7 (r 2 = 0.06, P = 0.01, n = 102) for group II, PN = 4.0 BW + 19.2 (r 2 = 0.19, P <0.0001, n = 101) for group III, PN = 11.6 BW + 66.4 (r 2 = 0.26, P <0.0001, n = 99) for all groups ( Fig. 13 View Figure 13 ).
Variation of paragnath number
Paragnath numbers in mature and immature adults collected from 8 localities are summarized in Table 2.
Karyotype
Diploid chromosome number was 28 (our unpublished data).
Reproduction
Reproductive swarming occurred at midnight just after high-tide during a few days throughout the spring tides in December at Kojima Bay, type locality ( Izuka, 1908). In the present study, swarming was observed just after high-tide at around 20:30 on 29 December 2001 at Nanaura in the Ariake Sea. In both cases, mature males and females swam near the surface above the tidal flats which are the habitat of immature worms, and shed sperm and eggs there.
Mature or nearly mature adults were collected from the sediment samples in the Omutagawa River on 4 January and 21 December 1999, and only juveniles were collected in March in Daijugarami , indicating that sexual reproduction occurs during late December to January in the Ariake Sea, and that the life span is 1 year. Large specimens collected from Inchon, Korea on 19 January 1999 contained immature oocytes (up to 130 Mm in diameter) or clusters of spermatocytes, indicating that sexual reproduction commences in late January or later in Korea .
Gamete morphology and development
The mature eggs (full-grown oocytes) were spherical and 180–210 Mm in diameter in the fixed specimens ( Fig. 14a View Figure 14 ). The mean diameter (± SD) of eggs contained in the body cavity of each swarming female was as follows: 187.1 ± 6.3 Mm (n = 25) for the lectotype collected from Kojima Bay, 184.2 ± 4.4 Mm (n = 14), 188.2 ± 4.8 Mm (n = 20) and 188.7 ± 7.7 Mm (n = 11) for three nontype specimens collected from the Ariake Sea. The oocytes contained 40–60 oil drops surrounding a germinal vesicle. The spermatozoa had a coneshaped acrosome at the tip of the head ( Fig. 14b View Figure 14 ). The sperm head was about 2.5 Mm wide, about 3.5 Mm long, and slightly angular at the base of acrosome.
After embryonic development (c. 2 days after fertilization), trochophore larvae swam actively, showing positive phototaxis; about 11 days after fertilization, most larvae with 4 or more setigers sank down to the estuary floor and became benthic ( Izuka, 1908).
Habitat
All specimens were collected from muddy tidal flats in estuaries with a wide tidal range (around 3 m or more in the mean tidal range in spring tides). Where the Omutagawa River flows into the Ariake Sea, this species occurred sympatrically with Hediste diadroma sp. nov.
Geographical distribution
The coasts of the Yellow Sea in Korea, the Seto Inland Sea and Ariake Sea in Japan ( Fig. 15 View Figure 15 ).
Remarks
Hediste japonica is easily distinguishable from all other Hediste species due to the presence of homogomph falcigers and the absence of heterogomph spinigers in the neuropodia, the presence of a digitate lobe at the tip of the postchaetal ligules throughout, and the fewer and larger paragnaths in group II.
Both the original and subsequent descriptions of this species ( Izuka, 1908, 1912) are incomplete, presenting little information on chaetal morphology and paragnath number. As for falcigers in the neuropodia, only a typical heterogomph form was described, while the homogomph form was not. These morphological descriptions were probably based on specimens of different species, e.g. H. diadroma sp. nov. or H. atoka sp. nov. (see below), which were both present in Izuka’s collections from the Sumida River and elsewhere ( Table 1). It is also strange that the diameter of the mature eggs was described as 140–160 Mm in the original description of H. japonica ( Izuka, 1908) . These values are characteristic of H. diadroma sp. nov. (see below), and different from those of the eggs remaining in the lectotype (180–200 Mm).
more species. Smith (1958) detected the presence of homogomph falcigers and an increase in the number of homogomph spinigers in the epitokes of H. japonica sensu stricto, which were probably collected from Kojima Bay (type locality). However, the presence of homogomph falcigers was incorrectly regarded as an epitoke-specific characteristic, probably because Smith (1958) examined atokous specimens of different species ( H. diadroma sp. nov. or H. atoka sp. nov.) for comparison with the epitokous ones of H. japonica .
Two populations of Nereis japonica sensu Khlebovich et al. (1982) , which were collected from the Yellow Sea, may belong to H. japonica sensu stricto because of the small number of paragnaths found in group II (7– 8 on the left side), and group IV (8–9 on the left side).













| T |
Tavera, Department of Geology and Geophysics |
| MNHN |
Museum National d'Histoire Naturelle |
| V |
Royal British Columbia Museum - Herbarium |
| VI |
Mykotektet, National Veterinary Institute |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hediste japonica
| Sato, Masanori & Nakashima, Akiyuki 2003 |
Hediste sp. 2
| Sato M 2001: 66 |
| Sato M 1999: 129 |
Nereis japonica
| Khlebovich VV & Komendantov AY & Yakovishina LA 1982: 1110 |
Hediste japonica: Imajima, 1972: 102–105
| Sato M 2001: 66 |
| Sato M 1999: 129 |
| Imajima M 1996: 139 |
| Imajima M 1972: 105 |
Neanthes japonica
| Smith RI 1958: 60 |
Nereis japonica
| Izuka A 1912: 163 |
| Izuka A 1908: 305 |