
Bothrioneurum, Stolc, 1886
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4497.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:11725C60-E463-4EB3-A96A-34CEF56923B8 |
|
DOI |
https://doi.org/10.5281/zenodo.5951342 |
|
persistent identifier |
https://treatment.plazi.org/id/5D62156B-FFDB-C84D-1AEC-D1FCFE246F32 |
|
treatment provided by |
Plazi |
|
scientific name |
Bothrioneurum |
| status |
|
The genus Bothrioneurum
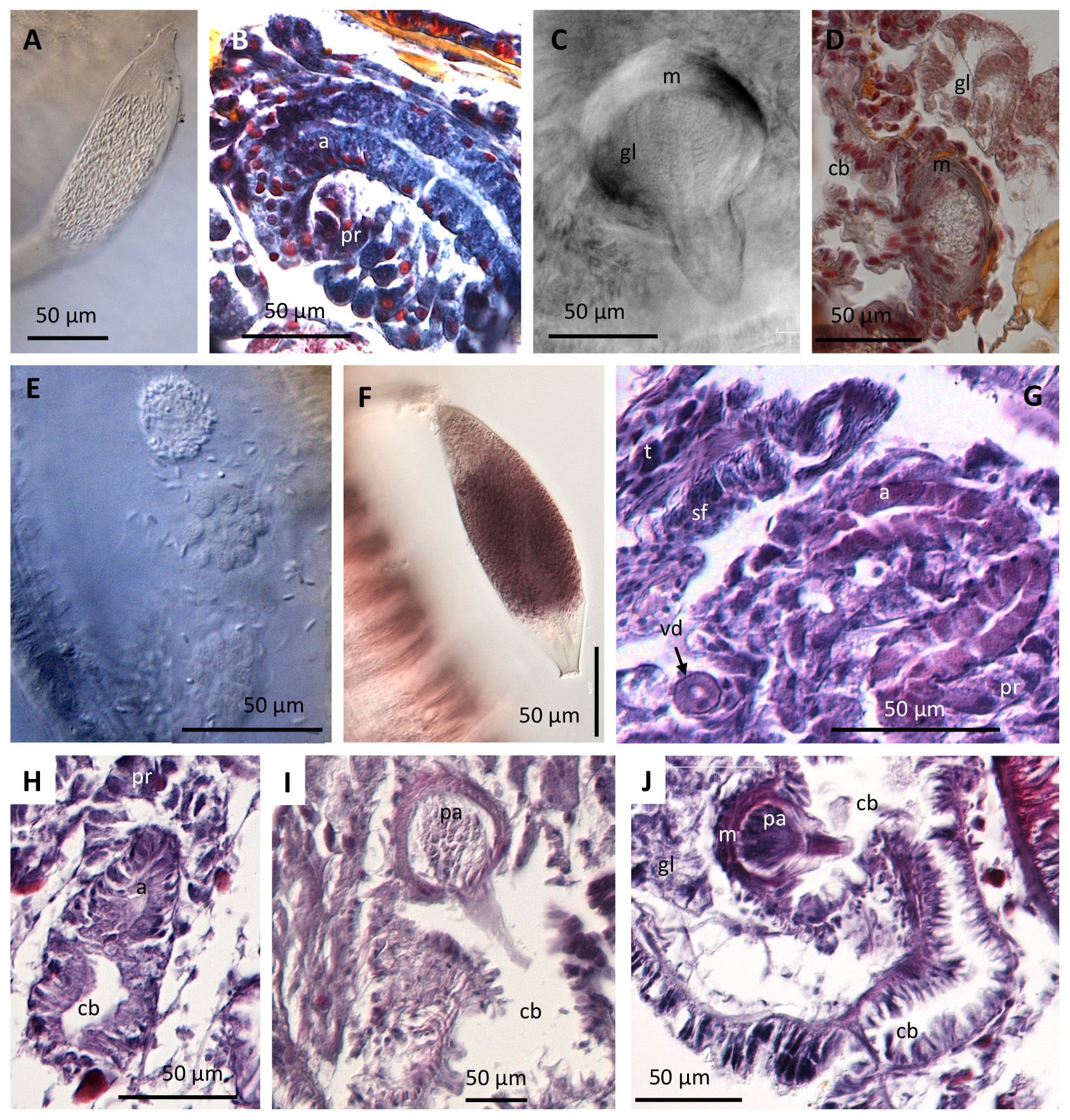
( Figure 2 View FIGURE 2 )
Spermathecae are absent in all known Bothrioneurum species, and asexual reproduction by fragmentation followed by regeneration of anterior segments seems to be common in the genus ( Brinkhurst 1971). Rhyacodrilines generally have the sequence of gonads starting in segment X, but the typical forward shift of the reproductive system in Bothrioneurum species seems associated with asexual reproduction ( Hrabě 1935; Timm 1979). Maturity appears to be regulated by temperature; in a laboratory culture, Timm (1997) observed that a few individuals of B. vejdovskyanum matured sexually, copulated (interpreted by the presence of spermatophores) and laid free cocoons only when water temperature sank to 13–15°C or lower in late autumn and winter, after many years of only asexual reproduction.
The spermatophores in Bothrioneurum spp. are formed by an oval, chitinous capsule with a solid stem firmly attached to the body wall ( Fig. 1A View FIGURE 1 ), and are not easily lost by handling the worm. The sperm cells of the examined specimens seem to have undergone little elongation of the nuclei ( Fig. 2A,E–G View FIGURE 2 ), thus appearing atypical among aquatic oligochaetes, but unfortunately we have not found any detailed report on sperm structure of Bothrioneurum species in the literature. Similar sperm is apparently shown in the illustration of B. pyrrhum Marcus 1942 (Plate X, Fig. 33), although flagella appear to develop on sperm of B. iris Beddard, 1901 ( Marcus 1942: Plate XI, Fig. 37). This possible lack of sperm elongation and flagella in some Bothrioneurum species requires further investigation, since it would be the first case not only for oligochaetes, but for the Clitellata (Marco Ferraguti, pers. comm. 30th July 2018).
The male duct includes a tubular atrium with diffuse prostate ( Fig. 2B,F,G View FIGURE 2 ), and a characteristic organ called the 'paratrium' ( Fig. 2C,D,I,J View FIGURE 2 ), which appears similar to glandular organs described in other taxa (see Uktena and Smithsonidrilus , below), and could act as a copulatory organ to attach the spermatophores to the partner ( Brinkhurst 1971). The term paratrium was attributed to Vejdovský by Beddard (1895), and probably refers to its position within the male duct and to its complex structure, including both muscular and glandular tissues. In most Bothrioneurum species, a bunch of gland cells enters a muscular bulb prolonged into a cone that protrudes into a deep male copulatory bursa ( Fig. 2C,D,I,J View FIGURE 2 ) (also referred to as copulatory chamber, copulatory sac, or 'eversible pseudopenis' in some descriptions). The body of the copulatory organ or paratrium is spherical to ovoid, surrounded by a muscular layer; a cluster of gland cells penetrates the muscle layer, forming a characteristic radial arrangement, and terminates in a conical ectal organ, which protrudes toward the male copulatory bursa ( Fig. 2C,J View FIGURE 2 ). The location of paratrium within the copulatory bursa differs among species; it is ectal in B. americanum Beddard, 1894 (see Černosvitov 1939: Fig 62), but ental in the others ( Table 1). A number of accessory glands can also be associated with the copulatory bursa in Bothrioneurum species, either ectal to the paratrium as in B. vejdovskyanum and B. brauni , or ental (as in B. americanum ).
The taxonomy of the genus Bothrioneurum has many unresolved issues, beyond the scope of this paper. Brinkhurst (1971: 540) suggested that all of the described species could be synonyms of B. vejdovskyanum , a species that is accepted as having a lot of variation, due to its predominant reproduction by fragmentation [see also Timm (1997) for a discussion on the taxonomic status of B. pyrrhum and B. iris ]. Although this genus requires a revision based on original or new collections, the number of "male pores" (openings of the copulatory bursae) and presence/absence of penial chaeta, together with the structure of the male duct, seem to support the differences between the species ( Table 1) until the necessary genetic analyses are performed. In the present study, we have had the opportunity to study specimens from an Argentinian population (Marchese's collection) that are most consistent with the description of B. brauni . The most relevant species-level taxonomic characters considered here are related to the male duct: size and position of the male funnel, relative length of the vas deferens to the atrium, presence of different sections in the male bursa, position of the paratrium, and presence of accessory glands. The new material has a large sperm funnel placed forward in the testicular segment ( Fig. 2G View FIGURE 2 ), the vas deferens is long and mostly contained within X; the atrium joins the copulatory bursa, forming a simple but abrupt transition ( Fig. 2H View FIGURE 2 ); the paratrium is in the ental part of the male bursa; male pores are paired (copulatory bursae open separately), and penial chaetae are absent. The ectal gland associated with the male pore described by Marcus (1949: Fig. 11) is visible only in one of the specimens.
The original description of B. brauni was based on only one mature individual, which had two pairs of testes and two pairs of ovaries, an atypical condition in the family Naididae , possibly related to asexual reproduction, whereas the specimens from Marchese's collection showed the normal gonadal arrangement (only one pair) for rhyacodrilines. Where present, the penial chaetae in the genus Bothrioneurum are of the rhyacodriline type, arranged fanwise, with the distal ends close together ( Rodriguez & Fend 2013). The only mention of penial chaetae for B. iris Beddard, 1901 was by Chen (1940), based on a single mature individual and suggested to be identical to B. vejdovskyanum (see Brinkhurst 1971: 541). B. vejdovskyanum , B. iris , B. schiemeri Timm, 1997 and B. grandisetosum Wang, Xie & Liang, 2000 have male pores opening within a single median chamber with a single external opening. In contrast, all American species have two distinct male pores (i.e., separate external openings of the copulatory bursae).
The paratrium is relatively similar in all Bothrioneurum species, but small differences in the thickness of the musculature, the presence of an apical gland cluster of cells on the paratrial bulb, or a cuticular sheath on the ectal cone that opens to the male bursa, have been considered diagnostic ( Table 1). In the examined material, it is not always easy to see the paratrial gland, which perhaps is more or less conspicuous depending on fixation or the state of maturity of the specimen. The new record of B. brauni in Argentina widens the distribution of the species in the Neotropical region.
Schmelz, 2000 was described based on immature worms. Bothrioneurum aequatorialis is not included as it is species inquirenda (after Brinkhurst 1971); sg: segment.
1 from the drawing in the original description
2 in VIII: Hrabĕ (1981), Fend's coll.; in IX: Ohtaka (2000); Timm (1979): references therein related to male opening in IX and XI
3 in Štolc (1888: Fig. 7)
4 in Timm (1979: Fig. 5.1)
5 in Ohtaka (2000: Fig. 1.K View FIGURE 1 )

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.