
Mimobdella japonica Blanchard, 1897: 94–95
|
publication ID |
https://doi.org/ 10.12782/sd.18.1.099 |
|
persistent identifier |
https://treatment.plazi.org/id/651587BB-FFE9-046E-FCAE-F9A0FE3FC6BD |
|
treatment provided by |
Felipe |
|
scientific name |
Mimobdella japonica Blanchard, 1897: 94–95 |
| status |
|
Mimobdella japonica Blanchard, 1897: 94–95 , pl. 6, figs 16, 17; Nakano 2011 a: 3–7, figs 1–4.
Diagnosis (amended from Nakano 2011). In life, dorsal surface ochre, ventral surface whitish ochre, paler than dorsal surface. Mid-body somites novem-annulate, generally c1=c2<b2<a2>c9=c10=d21=d22<c12. Clitellum in X c9 to XIII a2. Anus with 2–4 post-anal annuli. Post-crop caeca in pairs in XX c12 to XXII c1. Male gonopore in XI/ XII, female gonopore in XII/XIII, gonopores separated by 9 annuli (1 full somite). Sperm ducts reaching to XVI c1. Ovisacs reaching to XXIII c1, and then turning anteriorly.
Material examined. Two specimens collected from Okinawajima island , Okinawa Prefecture, Japan, by Masashi Sugimoto: KUZ Z227 View Materials , dissected, from Nago in spring of 2008, and KUZ Z229 View Materials , dissected, from Kin on 22 May 2011 . Two specimens collected from the underside of stones in a canal in a paddy field in Akina , Tatsugo , Amami-oshima island, Kagoshima Prefecture, Japan, by Takafumi Nakano on 28 April 2010: KUZ Z179 View Materials (alt. 0 m, 28°26′35″N, 129°33′36″E), dissected, and KUZ Z228 View Materials (alt. 0 m, 28°26′42″N, 129°33′38″E), dissected GoogleMaps .
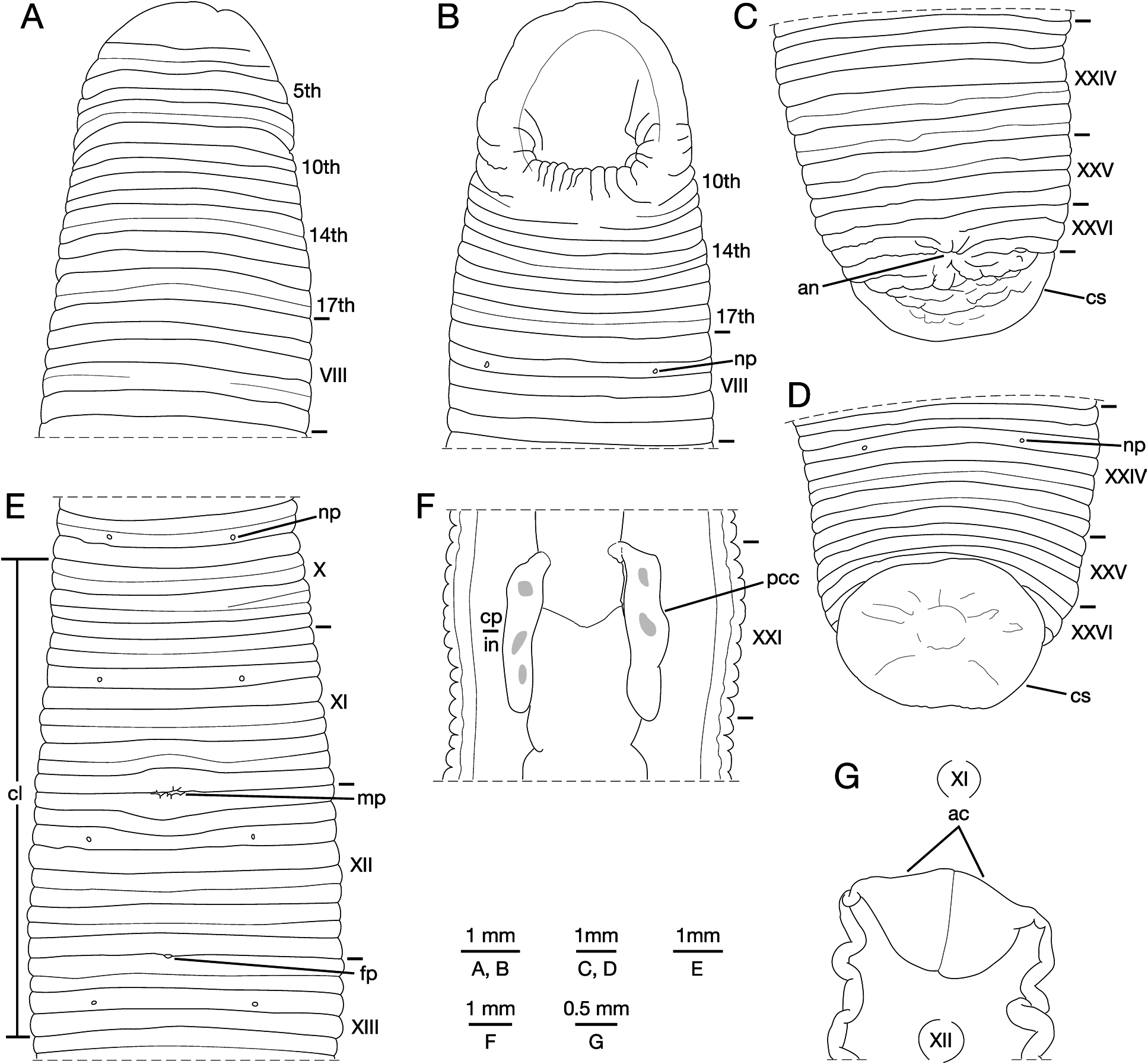
Description of specimens from the Ryukyu Islands. Body firm, muscular, elongated, with constant width in caudal direction, dorsoventrally depressed, maximum BL 106.88 (KUZ Z179), maximum BW 7.13 (KUZ Z227) ( Fig. 2A, B View Fig ). Caudal sucker situated ventrally, oval, its greater diameter smaller than BW ( Figs 2A, B View Fig , 4C, D View Fig ). In life, dorsal surface ochre, clitellum yellow ochre, ventral surface paler than dorsal surface, whitish ochre. Colour faded in preservative ( Fig. 3 View Fig ).
Annulation of somites I–VII unclear, comprising 14–17 annuli altogether, 1st annulus completely merged with prostomium, 7th–9th annuli forming posterior margin of oral sucker; KUZ Z179 with 15 annuli in this region, 12th and 15th annuli with obvious furrow; KUZ Z227 with 14 annuli, 9th and 11th annuli with slight dorsal furrow, 10th and 14th annuli with slight furrow; KUZ Z228 with 16 annuli, 12th, 13th and 16th annuli with slight furrow; KUZ Z229, comprising 17th annuli, 14th and 17th annuli with slight furrow, 7th and 8th annuli united on venter ( Fig. 4A, B View Fig ). Somite VIII quinquannulate, b1>b2<a2<b5>b6 (KUZ Z227, Z229) ( Fig. 4A, B View Fig ); or sexannulate, b1 (with slight furrow: c1, c2)>b2<a2<b5 (c9, c10)>c11>c12 (KUZ Z179, Z228). Somite IX quiquannulate, b1 (c1, c2)=b2 (c3, c4)<a2<b5 (c9, c10)=b6 (c11, c12) (KUZ Z229); sexannulate, b1 (c1, c2 dorsally)>b2=a2<b5>c11>c12 (KUZ Z227); or septannulate, c1=c2<b2<a2<b5=c11 (c9, c10)>c12 (d21, d22) (KUZ Z179, Z228). Somite X sexannulate, c1=c2<b2<a2<b5 (c9, c10)<b6 (d21, d22, c12) (KUZ Z227); octannulate, c1=c2=b2<a2<c9 (d17, d18)>c10 (d19, d20 ventrally)>c11=c12 (KUZ Z229) ( Fig. 4E View Fig ); or novem-annulate, c1=c2<b2<a2>c9=c10=d21= d22<c12 (KUZ Z179, Z228). Somite XI quiquannulate, b1 (c1, c2)>b2=a2<b5 (c9, c10)<b6 (d21, d22, c12) (KUZ Z227); or novem-annulate (KUZ Z179, Z228, Z229) ( Fig. 4E View Fig ). Somite XII sexannulate, b1 (c11, c12)>b2=a2<b5 (c9, c10)=c11 (d21, d22)>c12 (KUZ Z227); or novem-annulate (KUZ Z179, Z228, Z229) ( Fig. 4E View Fig ). Somite XIII septannulate, c1=c2=b2>a2<b5 (c9, c10)=c11 (d21, d22)>c12 (KUZ Z227); or novem-annulate (KUZ Z179, Z228, Z229). X c9 and XIII a2 respectively being first and last annuli of clitellum ( Fig. 4E View Fig ). Somites XIV–XXIII novem-annulate, or rarely somites XXII and XXIII respectively septannulate, c1=c2<b2<a2<b5 (c9, c10)=c11 (d21, d22)>c12, and quiquannulate, b1 (c1, c2)>b2<a2<b5 (c9, c10)<b6 (d21, d22, c12) (KUZ Z227). Somite XXIV quiquannulate, b1 (c1, c2)>b2<a2<b5 (c11, c12)=b6 (c11, c12) (KUZ Z227); sexannulate, c1=c2=b2<a2<b5 (c9, c10)=b6 (c11, c12) (KUZ Z229) ( Fig. 4C, D View Fig ); septannulate, c1=c2<b2<a2<b5 (c9, c10)>c11 (d21, d22)>c12 (KUZ Z228); or octannulate, c1=c2<b2<a2>c9=c10<c11>c12 (KUZ Z179). Annulation of somites XXV–XXVII comprising 7–9 annuli altogether, but hardly decidable; possibly XXV triannulate a1 (b1, b2)>a2<a3; XXVI triannulate, a1=a2=a3; and XXVII uni- (KUZ Z227), bi- (KUZ Z228), or triannulate (KUZ Z179, Z229); XXVI a1 (KUZ Z227, Z229), or a3 (KUZ Z179, Z228) being last complete annulus on venter; anus at XXVI a2/a3 with 2–4 post-anal annuli ( Fig. 4C, D View Fig ).
Eyes undectectable. Nephridiopores in 17 pairs in VIII– XXIV, situated ventrally at middle of b2 of each somite ( Fig. 4B, D, E View Fig ). Papillae numerous, minute, mainly 1 row on every annulus, and 2 or 3 rows on annuli with slight furrow(s).
Pharynx strepsilaematous, reaching to XIV d22/c12– XIV/XV, with 3 myognaths separated by triangular paragnaths, each myognath bearing 2 conical stylets arranged in tandem, parallel to body axis. Crop tubular, reaching to XXI b2–XXI d21; pair of post crop-caeca thin-walled in XX c12– XXI c2 to XXI c10–XXII c1 ( Fig. 4F View Fig ). Intestine tubular and acaecate, reaching to XXIII b1–XXIII c10/d21. Rectum tubular, thin-walled.
Male gonopore in XI/XII ( Fig. 4E View Fig ). Female gonopore in XII/XIII ( Fig. 4E View Fig ). Gonopores separated by 9 annuli (1 full somite). Testisacs multiple, in XV c12–XVI c2 to XXV a2– XXV a3, several testisacs on each side in each annulus. Sperm ducts in XI c12 to XV c10–XVI c1/c2, coiled, narrowing at junction with atrial cornu, then turning gradually inward toward atrial cornua without pre-atrial loop. Pair of atrial cornua curved laterad in XI c12 and XII c1 ( Fig. 4G View Fig ). Atrium short, muscular, globular, in XI c12 and XII c1. One pair of ovisacs long, thin-walled, slightly folded, tubular in XIII c1 to XXI c2/b2–XXIII c1/c2; right ovisac turned anteriorly in XXI c2/b2–XXIII c1/c2, then reaching to XX a2–XXII b2; left ovisac also turned anteriorly in XXI c2/b2–XXII c10, then reaching to XX a2–XXI d21, both ovisacs converging in XIII c1, directly descending to female gonopore.
Distribution and habitat. Known from Tasugo, Amami-oshima island, Japan, and lowland regions of Okinawajima island, Japan. Semi-aquatic species.
Genetic variation. The K 2P distance ( COI) between the two specimens of M . japonica from Okinawajima ( AB761393 View Materials , AB761395 View Materials ) and those from Amami-oshima ( AB679658 View Materials , AB761394 View Materials ) was 0 %. No difference was detected among the COI sequences obtained from these four specimens.
Remarks. Four specimens from the Ryukyu Islands were unambiguously identified as M. japonica based on their possession of the following characteristics: mid-body somites novem-annulate; clitellum in X c9 to XIII a2; male gonopore in XI/XII, female gonopore in XII/XIII, 9 annuli between gonopores; pair of post-crop caeca mainly in XXI; and paired sperm ducts reaching to XVI c1. Although the holotype of M. japonica possesses two post-anal annuli ( Nakano 2011), the Ryukyu specimens have two to four postanal annuli. In addition, the ovisacs of the present specimens are longer than those of the holotype of M. japonica . Therefore, the diagnosis of M. japonica is amended herein above to take these differences into account.
The COI sequence divergence among these four specimens is 0%. This low genetic diversity may indicate that the leeches have dispersed to these islands in a recent rapid range expansion, as have, e.g., species of Hirudo Linnaeus, 1758 in Europe ( Trontelj and Utevsky 2012), or that they have been introduced by human activities. The K2P distance among the specimens of M. japonica is abnormally low relative to that between Orobdella shimadae Nakano, 2011 and O. dolichopharynx Nakano, 2011 (8.5–11%, mean=9.7%, Nakano, unpub. data), which are also predaceous, but fully terrestrial leeches that are distributed in Okinawajima and Amami-oshima, respectively. Mimobdella japonica from Okinawajima has been found mainly in cultivated areas (Sugimoto, pers. comm.). Additionally, specimens of M. japonica from Amami-oshima have been collected only from paddy fields. Thus, M. japonica may have been introduced to either Okinawajima or Amami-oshima, or to both islands, via human activity. However, the origin of M. japonica in the Ryukyu Islands and the distributional range of this species elsewhere are still unclear. For example, large salifid leeches from Kyushu, Japan, are identified as Odontobdella blanchardi (Oka, 1910) , not M. japonica (Nakano, pers. obs.). Despite the fact that M. japonica can be regarded as a likely introduced species in the Ryukyu Islands according to its COI sequence divergence, its only confirmed localities are in these islands. Further faunal surveys are needed to confirm whether M. japonica is truly a rapidly dispersing alien species, and to fully reveal its range of distribution and biogeographical history.


| KUZ |
Zoological Collection of the Kyoto University |
| COI |
University of Coimbra Botany Department |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mimobdella japonica Blanchard, 1897: 94–95
| Nakano, Takafumi 2013 |
Mimobdella japonica
| Blanchard, R. 1897: 95 |