
Saccoglossus rhabdorhynchus, C. B. Cameron, C. Deland & T. H. Bullock, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.893894 |
|
DOI |
https://doi.org/10.5281/zenodo.6201829 |
|
persistent identifier |
https://treatment.plazi.org/id/68014E2E-FD39-FFA5-B8DD-FE37FBECFD50 |
|
treatment provided by |
Plazi |
|
scientific name |
Saccoglossus rhabdorhynchus |
| status |
sp. nov. |
Saccoglossus rhabdorhynchus View in CoL n. sp.
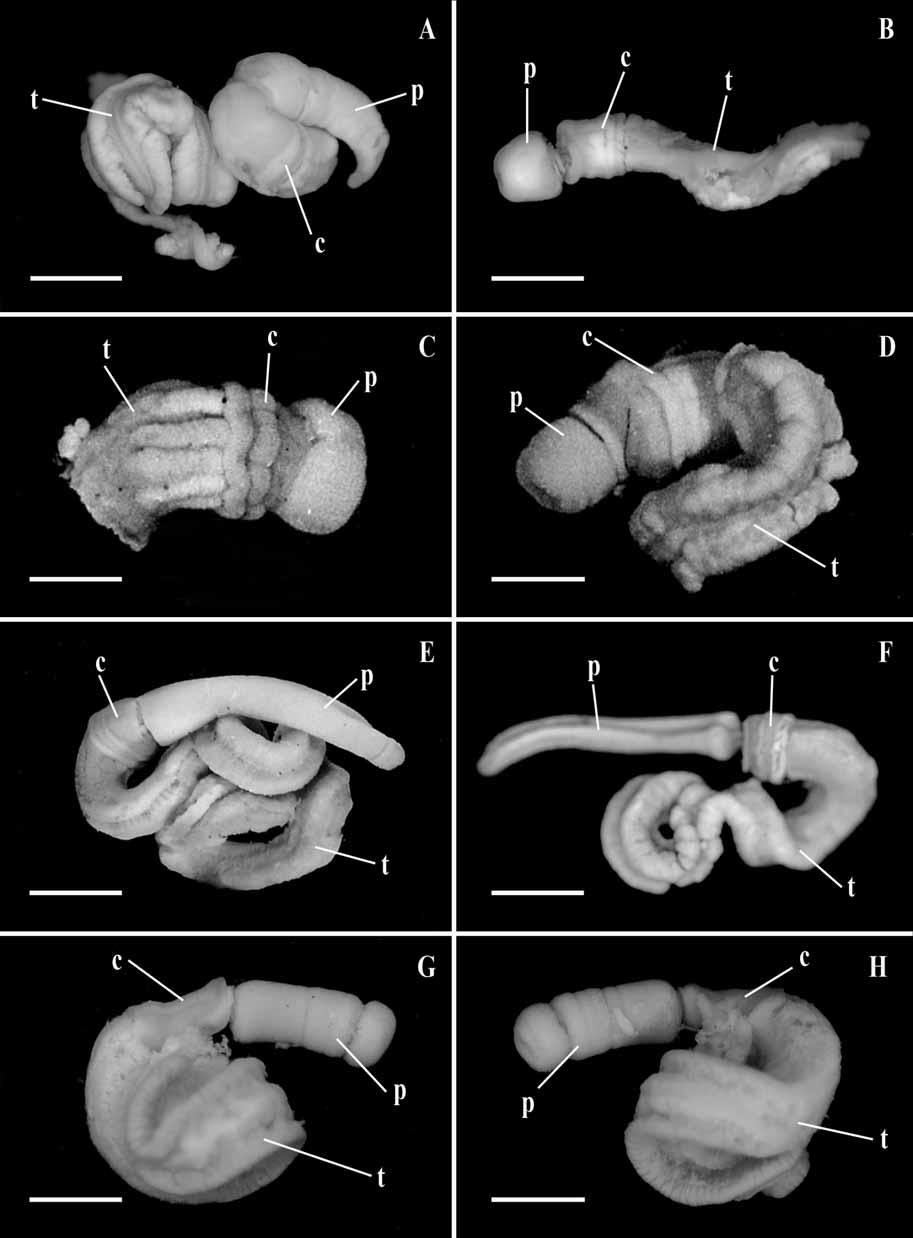
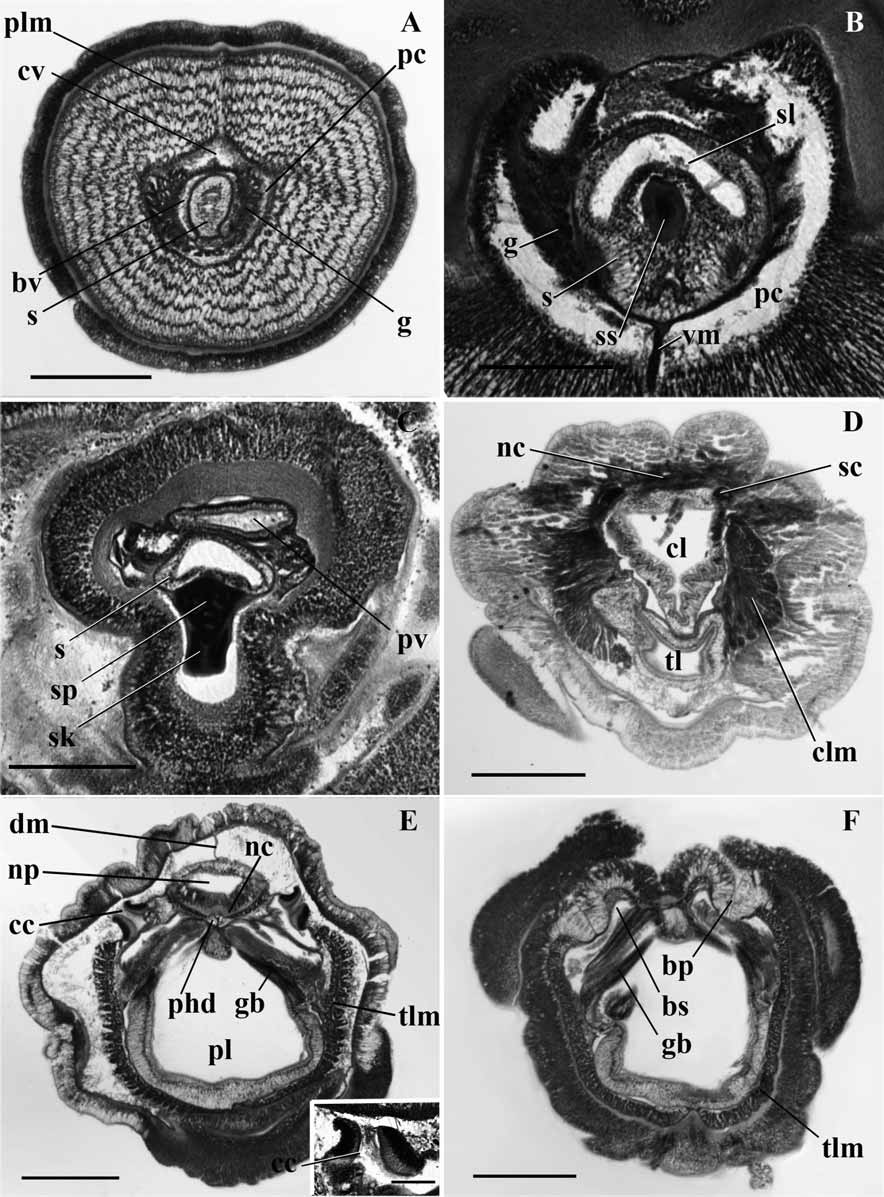
( Figs 1C,D View FIGURE 1 ; 3A–F)
Material examined. Six specimens were dredged in 1939 from a depth of 36 meters on rocky bottom off Half Moon Bay , California, (37° 12' N, 122° 30' W). The specimens were found in the surface debris on hard shale rock. Holotype: Accession no. NMNH 1132795 View Materials GoogleMaps . Paratype: Accession no. NMNH 1132796.
External features ( Figure 1 C, D View FIGURE 1 ): The total length is about 20 to 30 mm; the length of the proboscis is 2.5 mm, and the collar is 1.5 mm, after fixation. The collar is broader than long with two posterior annular thickenings. The posterior end of the collar is broader than the anterior. The length of the branchial region is about 4 mm. Openings of the branchial sacs are externally visible. The proboscis has a shallow middorsal longitudinal groove. The genital ridges are in the form of knob-like elevations on either side of the gill pores, extending posteriorly beyond the branchial region. The color in life is bright red.
Internal features ( Figure 3 View FIGURE 3 ): The nerve fiber layer of the proboscis is thickened middorsally along the shallow middorsal groove. The circular muscle fiber layer is quite conspicuous, being 4 or 5 fibers in thickness. Longitudinal muscle fibers are arranged concentrically, forming 8 or 9 rings. The proboscis cavity is narrow, extending nearly to the tip of the proboscis, completely filled anterior to the proboscis complex with irregular, granulated cells (connective tissue elements) limited all around by a layer of circular fibers. There is no dorsal septum and the ventral one extends only up to the ventral blind lumen of the stomochord ( Fig 3A View FIGURE 3 ). Only the left dorsal coelomic cavity is in communication with the median proboscis canal, which opens to the exterior by a left proboscis pore. The lumen of the stomochord is continuous extending nearly to the tip. The anterior part of the stomochord is slightly bent dorsal wards and this part shows only lacunae. The ventro lateral caeca of the stomochord lumen are united with one another and wide open to the primary lumen ( Fig. 3B View FIGURE 3 ). The cardiac vesicle extends ventrally over the sides of the stomochord. The glomerulus is present on the lateral sides of the stomochord and the halves are united anteriorly in front of the cardiac vesicle and the stomochord. The anterior face of the skeleton is projecting forward into a spine or obtuse projection ( Fig. 3B View FIGURE 3 ). The keel of the skeleton is quite distinct and deep anteriorly ( Fig. 3C View FIGURE 3 ); its cornua extend nearly to the posterior margin of the collar ( Fig. 3D View FIGURE 3 ).
Both dorsal and ventral mesenteries of the collar are incomplete, being confined to the posterior third or fourth of the collar. The collar canals are short and vertically oriented, the walls being thick, with nuclei arranged in many strata ( Fig. 3E View FIGURE 3 ). The perihaemal cavities extend up to the proboscis pore, separate from one another throughout. Peribuccal cavities are not distinguishable. The collar nerve cord is almost solid with only a few lacunae. It is continuous at both ends with epidermal invaginations ( Fig. 3E View FIGURE 3 ). No dorsal nerve roots are present. The stomochord opens into the buccal cavity in the anterior part of the collar. There is an invagination of the trunk gut in the posterior part of the collar that fuses posteriorly with the collar gut.
The ventral part of the pharynx is about a third of the total. The ciliated epithelium of the gill septa is thick with the nuclei in many strata. The ventral longitudinal musculature is heavily formed ( Fig. 3F View FIGURE 3 ). Gonads do not alternate with gill openings; only the lateral gonads are present.
Remarks. S. rhabdorhynchus is, as may be seen from the above description, similar to S. otagoensis in several characters, such as the extent of the skeletal cornua, the absence of the dorsal crest on the collar nerve cord, the absence of the ventral mesentery in the anterior part of the collar, the absence of lobed gonads, the origin of the genital ridges close to the collar and the arrangement in several strata of the nuclei in the ciliated branchial epithelium. One or more of these features are shared by certain other species of Saccoglossus , such as, S. horsti , S. shumaginensis , S. mereschkowskii , S. pusillus , S. kowalevskii , and S. ruber . While S. pusillus and S. ruber have only one character each in common with the species under consideration, S. horsti on the other hand is next only to S. otagoensis in its resemblance to the new species. Thus, they both have well developed ventral muscular ridges in the trunk; in both the genital ridges start near the collar; in both the gonads are not lobed; and finally in both the nuclei of the ciliated branchial epithelium are in several strata ( Table 1).
Although S. rhabdorhynchus more or less resembles certain other species of Saccoglossus , it exhibits some features that are shared by no other species of the genus. The complete filling up of the proboscis coelom by irregular granulated cells, the possession of a dorsal glomerulus and the complete separation of the perihaemal cavities through their entire length are characteristic of this species of Saccoglossus . The type of substratum from which this species was collected, hard shale rock, is also rather unexpected. Saxipendium coronatum lives upon rock ( Woodwick & Sensenbaugh, 1985) but this type of habitat has no parallel among the known species of Saccoglossus and is to be considered a unique feature of this species, unless in fact the debris on the shale amounts to a considerable layer of muddy sand.
Etymology. The trivial name of this species is derived from the fact that the proboscis coelom is solidly filled with granulated cells unlike that in all other known species of the genus.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |