
Bathyvermilia gregrousei, Kupriyanova, Elena K. & Ippolitov, Alexei P., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4044.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:4ABB85F0-D989-4702-9962-BA6E4D26BDE2 |
|
DOI |
https://doi.org/10.5281/zenodo.5324184 |
|
persistent identifier |
https://treatment.plazi.org/id/741287F4-FFE9-8C26-FF25-F9E493D521F9 |
|
treatment provided by |
Plazi |
|
scientific name |
Bathyvermilia gregrousei |
| status |
sp. nov. |
Bathyvermilia gregrousei View in CoL sp. nov.
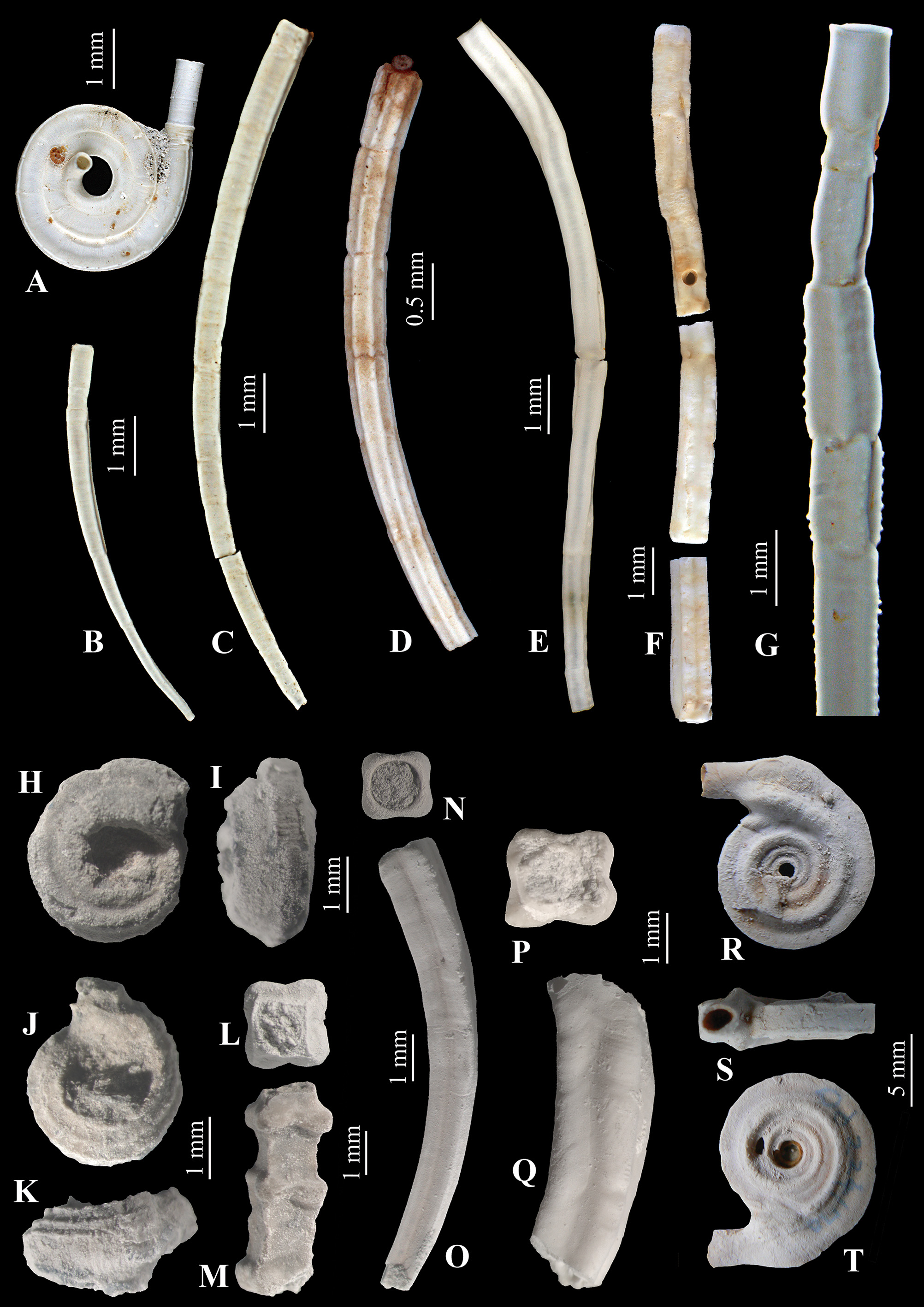
Figures 1 View FIGURE 1 E, 8–10
?"tube de serpulien".— Fauvel 1909: 74–75.— Fauvel 1914: 338–339, pl. 29, fig. 2–6.
Spirodiscus grimaldii View in CoL .— Hartman & Fauchald 1971: 183 [in part, R/V ATLANTIS-II, St. A119].
Material examined. Prince of Monaco, cruise 1896: Azores; St. 663, 27.6.1896, 37°28'30"N, 25°31'45"W, 1732 m (empty tubes, MOM 18 2642); St. 698, 18.7.1896, 39°11'N, 30°44'40"E, 1846 m (empty tubes, MOM 18 0475).
R/V JEAN CHARCOT, cruise INCAL, SW Ireland - off Brittany: St. 2.1-Pr34-WS1, 30.7.1976, 50°19.4'N, 13°08.1'W, 2539–2550 m (holotype MNHN POLY TYPE 1567,> 50 paratypes MNHN POLY TYPE 1568, 1 paratype prepared for SEM AM W.46401,> 100 paratypes AM W.46400, 3 tubes of this set prepared for SEM PIN 5485/13, PIN 5485/35, PIN 5485/36 and X-ray diffraction analysis sample #1,> 50 paratypes NBCL ZMA V.P o l 5550,> 50 paratypes SMF 23990,> 50 paratypes NHMUK ANEA 2015. 937–946,>50 paratypes LACM-AHF Poly 7022,> 50 paratypes USNM 1283059); St. 2.1-Pr29-CP9, 27.7.1976, 50°15.4'N, 13°15.8'W, 2659–2691 m (> 30 spec. MNHN PNT 51,>50 spec. AM W.46402,>30 spec. NBCL ZMA V.P o l 5554,>30 spec. SMF 23991, 12 spec. NHM UK ANEA 2015.955–964,> 30 spec. LACM-AHF Poly 7024,> 30 spec. USNM 1283062); St. 2.2- Pr37-CP10, 31.7.1976, 48°25.5'N, 15°10.7'W, 4823 m (5 spec. broken in pieces SMF 23992); St. 2.2-Pr39-DS11, 1.8.1976, 48°18.8'N, 15°11.5'W, 4823 m (1 spec. SMF 23933); St. 2.4-Pr48-OS3, 4.8.1976, 46°02.9'N, 10°18.7.1'W, 4798 m (tube fragments SMF 23994). Same, cruise BIACORES, Azores: St. 92, 17.10.1971, 39°03.5'N, 28°27.5'W, 2450 m (fragments of 1 empty tube MNHN PNT 48); St. 165, 1.11.1971, 37°33'N, 25°58'W, 2050–2085 m (2 spec. in tubes MNHN PNT 47); St. 171, 1.11.1971, 37°58.5'N, 26°07'W, 3215 m (9 spec. SMF 23977); St. 173, 2.11.1971, 37°57'N, 26°08'W, 3225 m (5 spec. NBCL ZMA V. P o l 5552, 5 spec. USNM 1283060, 5 spec. LACM-AHF Poly 7023); St. 174, 2.11.1971, 38°06'N, 26°15'W, 3050–3100 m (1 spec. in fragmented tube, SMF 23996). Same, cruise BIOGAS 11, off Brittany: St. CP37, 11.10.1981, 47°33.8'N, 8°39.2'W, 2175 m (2 spec. SMF 23999).
R/V DISCOVERY, cruise 105 (biology), off Brittany: St. 10112#1, 9.9.1979, 50°25.0'N, 13°19.1'W, 2640– 2660 m (7 spec. in tubes SMF 24000); St. 10112#3, 9.9.1979, 50°19.1'N, 13°25.8'W, 2740–2755 m (1 spec. and tube fragment MNHN PNT 49).
R/V CHALLENGER, cruise 5/82 (biology cruise 514), off Brittany: St. 51416: 31.3.1982, 50°16.8'N, 13°31.4'W, 2770–2780 m (5 spec. MNHM PNT 50, 5 spec. USNM 1283061, 5 spec. NHM UK ANEA 2015.950– 954, 3 spec. AM W.46405, 5 spec. NBCL ZMA V.P o l 5553).
R/V LE SUROIT, cruise EPI 1, off Brittany: St. CP39, 30.3.1984, 47°32.0'N, 8°38.4'W, 2100 m (1 spec. SMF 23998).
R/V CRYOS, cruise ABYPLAINE, North-East Atlantic: St. 2-DS1, 17.5.1981, 37°18'N, 15°33'W, 4260–4450 m (tube fragments SMF 23997); St. 8-CP11, 30.5.1981, 34°06.1'N, 17°06.3'W, 4270 m (5 spec. NBCL ZMA V.P o l 5551); St. 10-DS10, 11.6.1981, 42°51.2'N, 15°55.3'W, 4270–4360 m (3 spec. AM W.46403); St. 10-CP18, 11.6.1981, 42°52.3'N, 15°53.1'W, 4330 m (3 spec. NHM UK ANEA 2015. 947–949).
R/V ATLANTIS-II, North-West Atlantic: St. A119, 19.8.1966, 32°15.8' to 32°16.1'N, 64°31.6' to 64°32.6'W, 2095–2223 m (7 spec. LACM-AHF); St. A155, 00°03'S, 27°48'W, 3730–3783 m (8 spec., 1 spec. prepared for SEM, LACM-AHF).
R/V MARION DUFRESNE, cruise MD 50, Amsterdam Island: St. 5-DC34, 13.7.1986, 37°40.33'S, 77°30.50'E, 2200 m (7 spec. MNHN PNT 52, 7 spec. AM W.46404, 7 spec. NBCL ZMA V.Pol. 5555, 7 spec. SMF 24001, 7 spec. NHM UK ANEA 2015.965–971, 7 spec. LACM-AHF Poly 7025, 7 spec. USNM 1283063).
R/V VITYAZ, Pacific Ocean: St. 3243, 11.5.1955, 39°43.8'N, 159°48.0'E, 5542 m (1 spec. SIO); St. 4191, 8.12.1958, 40°22.5'N, 135°49.9'W, 4472–5072 m (2 spec. SIO); St. 4265, 13.01.1959, 24°57.6'N, 113°24.8'W, 3315–3340 m (2 spec. SIO); St. 4279, 19.1.1959, 19°46'N, 120°17.4'W, 4104 m (1 spec. SIO); St. 4281, 21.01.1959, 20°01.3'N, 121°59.6' W, 4370 m (1 spec. SIO); St. 4370, 3.3.1959, 26°04.2'N, 153°49.3'W, 6127– 6107 m (1 tube SIO); St. 5937, 0°20.2'N, 179°52'W, 5480 m (2 spec. SIO); St. 6298-56, 22°41.9'N, 160°50.8'W, 4270–4350 m (2 tube pieces SIO).
(?) R/V VITYAZ-2, Atlantic Ocean: St. 79, 34°54.3’N, 45°39’W, 4440 m (1 tube studied with SEM, PIN 5485/14 and X-ray diffraction analysis, sample # 2).
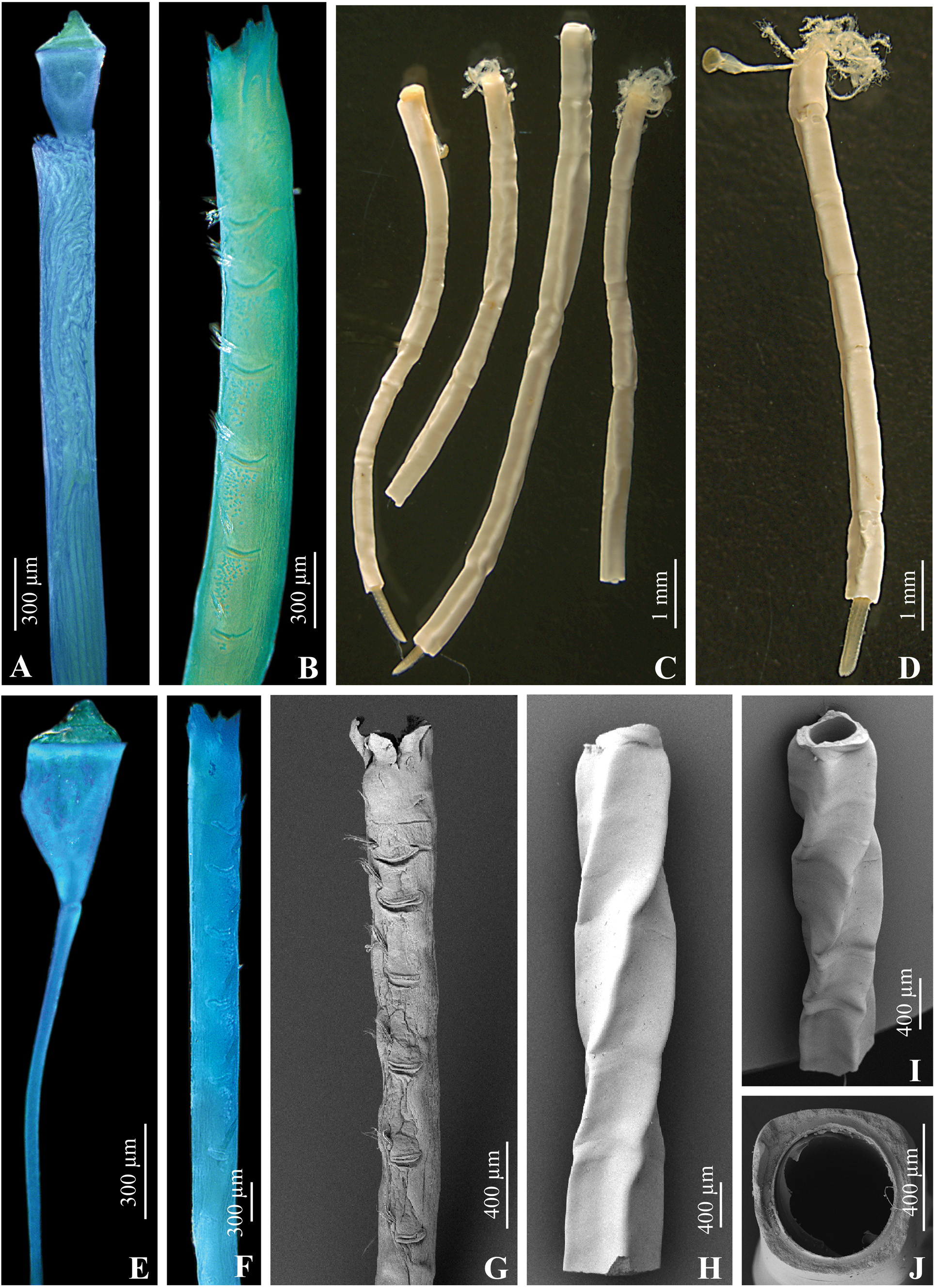
Description. Tube: white, opaque, ostensibly free, with shiny surface, open at both ends, straight or slightly curved, slowly increasing in diameter, slightly twisted spirally ( Fig. 8 View FIGURE 8 C, D, H, I). Cross-section tetragonal ( Fig. 8 View FIGURE 8 J). Sides of tube slightly concave, but cross-section becomes almost circular anteriorly ( Fig. 8 View FIGURE 8 J). Some tubes showing growth stops marked by tiny constrictions, not peristomes, accompanied by sudden turns around growth axis up to 45° ( Fig. 8 View FIGURE 8 C, D), some tubes can be slightly twisted ( Fig. 8 View FIGURE 8 H, I). Wall thickness varying between tube sides, with one side (lateral?) being significantly thinner ( Fig. 9 View FIGURE 9 F), making tube cross-section bilaterally symmetrical (see Size as well).
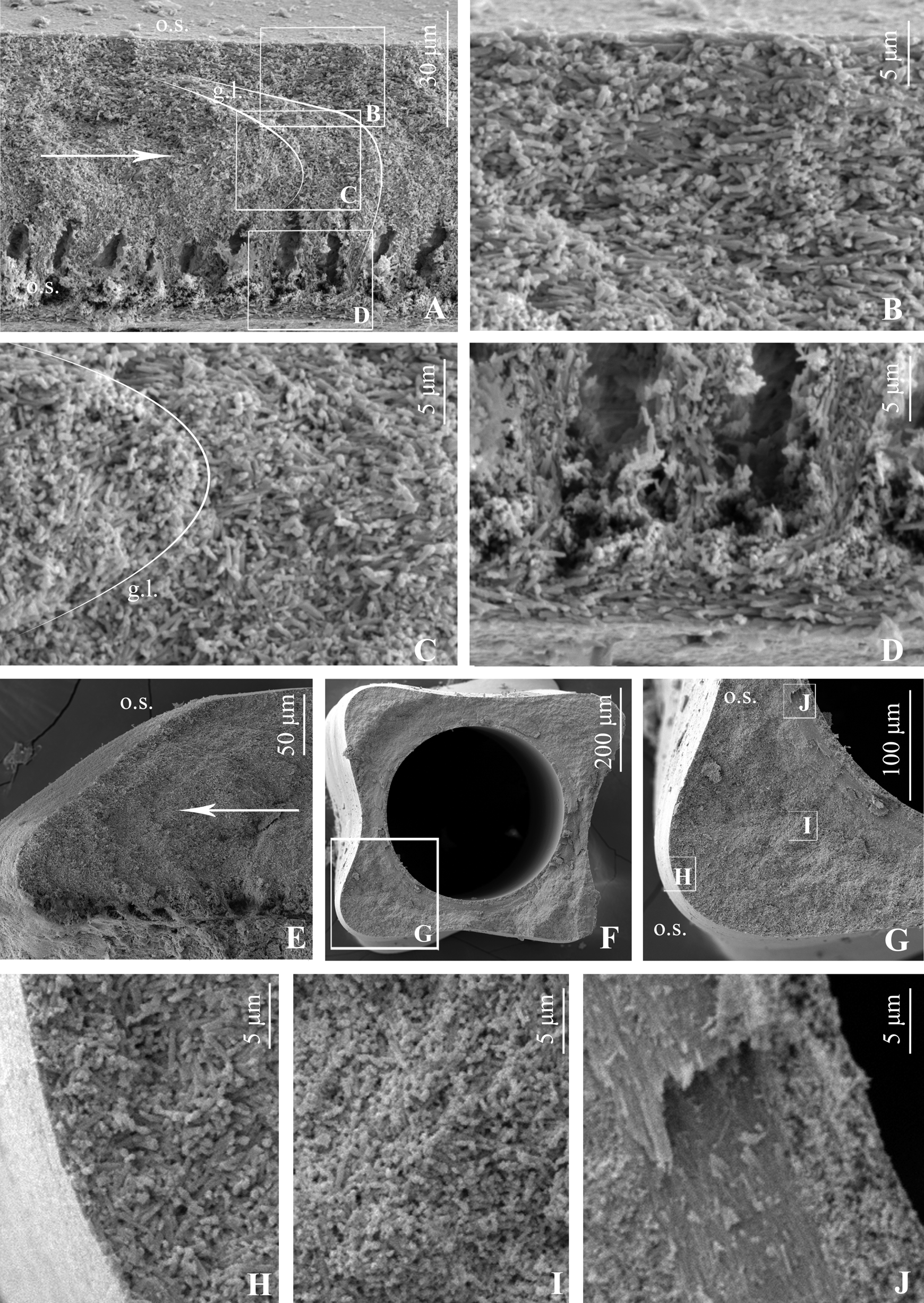
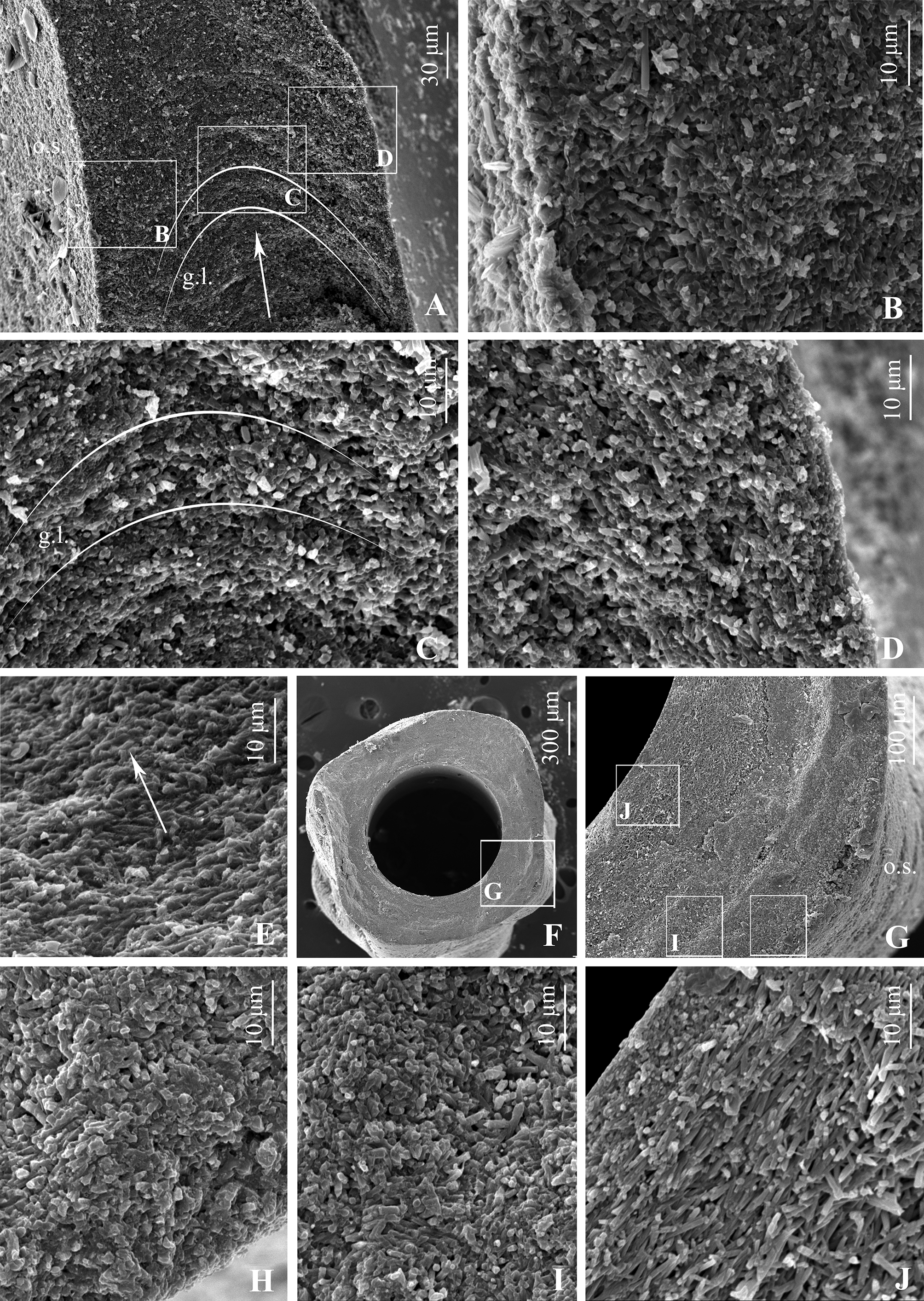
Tube ultrastructure: wall unilayered, with irregularly oriented prismatic (IOP) structure consisting mostly of elongated cigar-shaped crystals, but appearance and orientation of crystals throughout the wall not uniform. Inner part thin (1/10–1/15 of wall, Fig. 9 View FIGURE 9 D), but clearly distinct; consisting of consolidated rice grain-shaped crystals oriented more or less along growth direction by their axes. Length of crystals about 3–4 µm, diameter 0.5 µm. Middle wall part separated from inner layer by characteristic zone with numerous elongated “cavities”, positioned along growth lamellae of tube wall ( Fig. 9 View FIGURE 9 A, D, E). These cavities entirely surrounding lumen also visible in transverse sections ( Fig. 9 View FIGURE 9 G, J). Middle part of tube wall ( Fig. 9 View FIGURE 9 C, I) consisting of unoriented crystals similar in shape and size to crystals of inner and outer parts. Outer wall part ( Fig. 9 View FIGURE 9 B, H) composed of elongated crystals similar to crystals of inner layer, also oriented loosely parallel to tube wall. All variations of wall thickness produced by middle and outer wall parts ( Fig. 9 View FIGURE 9 G), thickness of inner part and “cavities” zone being stable. Parabolic growth lamellae indistinct, but visible ( Fig. 9 View FIGURE 9 A, C) in sections.
Tube mineralogy: sample # 1: 20% calcite (I calc=9), 80% aragonite (I arag=37); sample # 2: 55% calcite (I calc=56) and 45% aragonite (I arag=44).
Radiolar crown: 4–6 pairs of radioles arranged in semicircles to pectinately, not joined by inter-radiolar membrane. Radiolar eyes not visible in preserved material. Long naked tips of radioles absent.
Peduncle: inserted slightly in front between 1st and 2nd dorsal radioles, about same thickness as radioles, smooth (no pinnules), slightly longer than radioles ( Fig. 8 View FIGURE 8 E).
Operculum: elongated, covered with convex yellow-brown chitinous cap, sometime with a knob in the centre ( Fig. 8 View FIGURE 8 A, E); opercular bulb gradually narrowing towards peduncle, but separated by shallow conspicuous constriction.
Collar and thoracic membranes: collar short, subdivided into 2 latero-dorsal and 1 ventral lobe, the latter slightly incised. Thoracic membranes reaching up to 2nd chaetiger, about same width throughout.
Thorax: with 7 thoracic chaetigers, 6 of which uncinigerous ( Fig. 8 View FIGURE 8 B, F, G). Tori widely separated, first and second slightly closer to each other. Thoracic tori of similar size along thorax. Collar chaetae as few short capillaries (Fig. 10B); other thoracic chaetae limbate, with short slightly bent distal blades; Apomatus chaetae present (Fig. 10C). Uncini saw-shaped with 9–11 teeth in profile view, dental formula P:1:1:1:1:1:1:1:1:1 (Fig. 10D); anterior peg pointed.
Abdomen: with up to 45 segments. Uncini similar to thoracic ones, but rasp-shaped, with 3–4 rows, dental formula P:3:3:4:4:4:4:3:3:3 (Fig. 10F). Chaetae flat triangular blade with rounded denticles, only slightly longer on posterior segments (Fig. 10K); each chaetiger usually with a single chaeta. No achaetigerous zone between thorax and abdomen. Posterior glandular pad absent.
Size: total body length up to 15 mm, including up to 11 mm long radioles, width of thorax up to 0.4 mm. Maximum tube fragment length observed 35 mm (tubes incomplete). External tube diameter up to 0.67 mm with corresponding lumen diameter up to 0.45 mm. Thickness of tube wall in between angular margins about 1/7th of outer diameter at thinner sides, and ~1/6th at thicker sides.
Distribution. North Atlantic, low bathyal to abyssal depths (1732–4823 m), Indian Ocean (Amsterdam Island), 2200 m, Pacific Ocean, 3315–6127 m.
Etymology. The species is named after Professor Greg Rouse (Scripps Institution of Oceanography, USA) who made important contributions to serpulid phylogeny and deep-sea biology.
Remarks. This species apparently was first characterised by Fauvel (1909, 1914) from empty tubes only: “quadrangular in cross-section and thereby similar to Spirodiscus grimaldii but straight”. Similar tubes from the Indian Ocean (near Amsterdam Island, 2200 m) were found co-occurring with typical coiled S. grimaldii . These Indian Ocean specimens of B. gregrousei sp. nov. also have the distal parts of their tubes smooth and circular in cross-section. Occasionally these parts can be proportionally very long.
The tubes of Bathyvermilia gregrousei sp. nov. resemble quadrangular tusk-shaped tubes of Bathyditrupa hovei in size and tube diameter (compare Fig. 1 View FIGURE 1 B, C with E). However, tubes of B. hovei have a very regular slightly curved shape, a quadrangular tube cross-section with tube edges forming straight angles throughout and distal parts of the tube never become circular in cross-section. The tubes of Bathyvermilia gregrousei sp. nov., although more or less straight and quadrangular in cross-section, have more variable and less regular shapes ( Fig. 11 View FIGURE 11 C, D), normally slightly spirally twisted ( Fig. 11 View FIGURE 11 H, I), and distal tube parts often become smooth and circular in cross-section. However, the morphology of the animals removed from their tubes is very different.
Known ultrastructures for Bathyvermilia View in CoL cover three species— B. islandica Sanfilippo, 2001 View in CoL (tube structure is illustrated in the original description), B. langerhansi ( Fauvel, 1909) View in CoL figured by Vinn (2008) and Vinn et al. (2008) and Bathyvermilia challengeri Zibrowius, 1973 View in CoL (figured in Kupriyanova et al. 2014), thus allowing intrageneric comparison. All three species have tubes with well-developed homogeneous angular crystal ultrastructure (HAC) at least in the outer layer. This highly specialized structure has nothing to do with the IOP ultrastructure described herein for B. gregrousei View in CoL sp. nov. IOP ultrastructure constituting the inner layer of B. islandica View in CoL and the entire tube of B. gregrousei View in CoL sp. nov. is widely distributed among serpulids (Vinn et al. 2008, table 2) and seems to be less specialized, which suggests that HAC ultrastructure may be an apomorphy inside a certain group of Bathyvermilia View in CoL species.
Tube ultrastructure of B. gregrousei View in CoL sp. nov. in general is similar to those described for Bathyditrupa View in CoL and Spirodiscus , being also composed of elongated crystals similar in size and shape. However, the inner part of the wall of B. gregrousei View in CoL sp. nov. is made of larger elongated, not smaller isometric crystals. Moreover, an outer layer with SOIOP structure representing crystal “bundles” oriented subparallel to the tube surface and growth direction, characteristic for Spirodiscus and Bathyditrupa View in CoL , is absent in B. gregrousei View in CoL . At the same time, unilayered tube walls built of more or less chaotically oriented elongated crystals are not unique to Bathyvermilia View in CoL but known for a wide range of serpulid taxa (see Vinn et al. 2008). While making comparisons based on descriptions only is difficult, among figured ultrastructures the most similar one is known for Protis hydrothermica View in CoL (see ten Hove & Zibrowius 1986, fig. 4a, b) that seems to be relatively close to Bathyvermilia (Kupriyanova & Nishi 2010) View in CoL . Very diagnostic for the B. gregrousei View in CoL sp. nov. are microcavities surrounding the lumen and observed in all three specimens studied with SEM. Although cavities of various morphology are known in some taxa (e.g., Bianchi 1981: fig. 27b, c, 32a– c), such tiny ones surrounding the entire lumen were not previously described for any other serpulid.
| MOM |
Musee Oceanographique Monaco |
| MNHN |
Museum National d'Histoire Naturelle |
| PIN |
Paleontological Institute, Russian Academy of Sciences |
| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
| NHMUK |
Natural History Museum, London |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| MNHM |
John May Museum of Natural History |
| SIO |
Scripps Institution of Oceanography |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bathyvermilia gregrousei
| Kupriyanova, Elena K. & Ippolitov, Alexei P. 2015 |
Spirodiscus grimaldii
| Hartman 1971: 183 |