
Octomagelona Aguirrezabalaga, Ceberio and Fiege,2001
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlad069 |
|
publication LSID |
lsid:zoobank.org:pub:65B60DD3-64C9-4262-B7B2-74DA4D3D889F |
|
DOI |
https://doi.org/10.5281/zenodo.10497556 |
|
persistent identifier |
https://treatment.plazi.org/id/743EE917-FFE5-FF98-FCD3-273C1CB2F982 |
|
treatment provided by |
Plazi |
|
scientific name |
Octomagelona Aguirrezabalaga, Ceberio and Fiege,2001 |
| status |
|
Genus Octomagelona Aguirrezabalaga, Ceberio and Fiege,2001 View in CoL
Type species: Octomagelona bizkaiensis Aguirrezabalaga et al., 2001 . Gender: female.
Diagnosis: [Aguirrezabalaga et al. (2001) and Mortimer (2019), emended.] Body divided into thoracic region comprising achaetous first segment and eight chaetigers, and abdominal region with unknown number of segments. Prostomium large, flaưened, wider than long with small frontal horns. Parapodia biramous, with noto- and neuropodial lateral lamellae filiform if present, superior dorsal lobes [ DML of Jones (1963)] absent. Branchiae absent. Thoracic chaetae long, limbate capillaries, abdominal chaetae tridentate hooded hooks. Posterior part and pygidium unknown. Lateral pouches not observed.
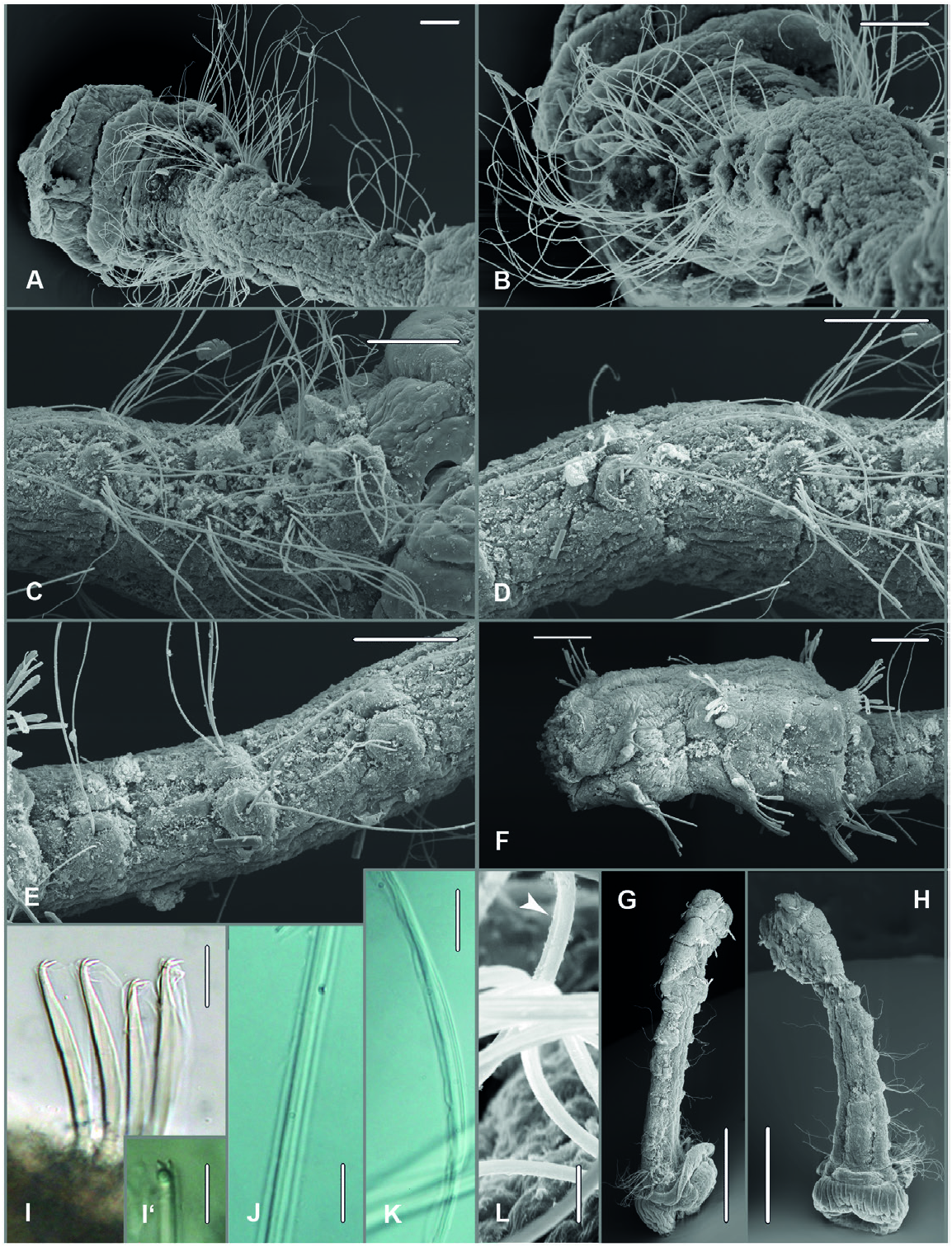
Remarks: Members of the genus Octomagelona are characterized by the presence of eight instead of nine thoracic chaetigers in contrast to all species of Magelona Müller, 1858 , the only other genus in Magelonidae . Instead of being a kind of intermediate segment sometimes equipped with special chaetae, chaetiger 9 represents the first abdominal segment in Octomagelona . The generic diagnoses provided by Aguirrezabalaga et al. (2001) and Mortimer (2019) has been slightly emended due to abdominal hooded hooks being tridentate in both species belonging to the genus Octomagelona , i.e. O. bizkaiensis and O. borowskii sp. nov.. Mortimer et al. (2021), considered Octomagelona a junior synonym of Magelona following their phylogenetic analysis based on morphological characters. We agree with these authors that the ‘magelonid-like body regionation’ is an important synapomorphy for Magelonidae ( Mortimer et al. 2021: 67 and abstract) but we regard the two different character states for the thoracic body region, i.e. the presence of eight versus nine thoracic chaetigers, as an easy to observe and taxonomically sufficient character to distinguish between the two genera. Therefore, we prefer to maintain Octomagelona as a valid genus for the time being and hope that studies of genetic markers will add significant information to characterize the two genera. Additionally, SEM and LM studies of thoracic chaetae in O. borowskii sp. nov., as well as the paratype of O. bizkaiensis , revealed that thoracic capillary chaetae are, in fact, unilimbate in O. bizkaiensis ( Fig. 12L View Figure 12 ) as stated in the original description (Aguirrezabalaga et al. 2001) and not bilimbate as coded by Mortimer et al. (2021: table S1, coding matrix character 31). In specimens of Octomagelona borowskii sp. nov. collected from the Peru Basin, thoracic capillaries were partly found to be bilimbate ( Figure 12J View Figure 12 ) as reported for the majority of Magelona species by Brasil (2003: table 5, coding matrix character 19) and Mortimer et al. (2021). But irregularly bilimbate chaetae are also present, i.e. with limbus irregularly indented ( Figure 12K View Figure 12 ) as defined by Brasil (2003: fig. 28B, C). Limbation of chaetae needs to be judged at high magifications and it remains to be evaluated whether irregularly indented limbation represents a natural character state or an artefact due to the effect of fixative or mechanical treatment during sampling.
Octomagelona borowskii sp. nov. Fiege, Knebelsberger and Meissner
( Figs 12A–F, I–K View Figure 12 )
Magelona sp. A Borowski, 1996: 54–55.
Type material: Holotype. SE Pacific Ocean, Peru Basin ( DISCOL area), SO 242-1 ( JPIO-DISCOL 1 ) stn 117-7 EBS, 4154 m depth, 19 Aug 2015, one af (11 chaetigers, length 3.0 mm, width at posterior end of prostomium 1.1 mm) ( SMF 30508 View Materials ).
Paratypes: SE Pacific Ocean, Peru Basin ( DISCOL area), SO 64 ( DISCOL 2 ) : one af, stn 1272 BC 1, 4159 m, 13 Feb 1989 ( SMF 30518 View Materials ) ; one af, stn 1302 BC 1, 4136 m, 22 Mar 1989 ( ZMH-P 30452 ) ; one af, stn 1379 BC 1, 4152 m, 9 Nov 1989 ( SMF 30520 View Materials ) ; one af + tentacle, stn 1388 BC 1, 4167 m, 13 Sep 1989 ( SMF 30521 View Materials ) ; one af, stn 1419 BC 1, 4156 m, 19 Sep 1989 ( SMF 30522 View Materials ) . SO 77 ( DISCOL 3 ) : one af, stn 1471 BC 1, 4152 m, 13 Feb 1992 ( ZMH-P 30451 ) ; one af, stn 1476 BC 1, 4174 m, 14 Feb 1992 ( SMF 30528 View Materials , SEM stub 1320) ; one af, stn 1492 BC 1, 4163 m, 21 Feb 1992 ( SMF 30530 View Materials ) ; all originally identified by C. Borowski as Magelona sp. A .
Measurements for largest paratype (SMF 30530): length 2.4 mm, width 0.8 mm (excl. parapodia) at posterior end of prostomium, 11 chaetigers.
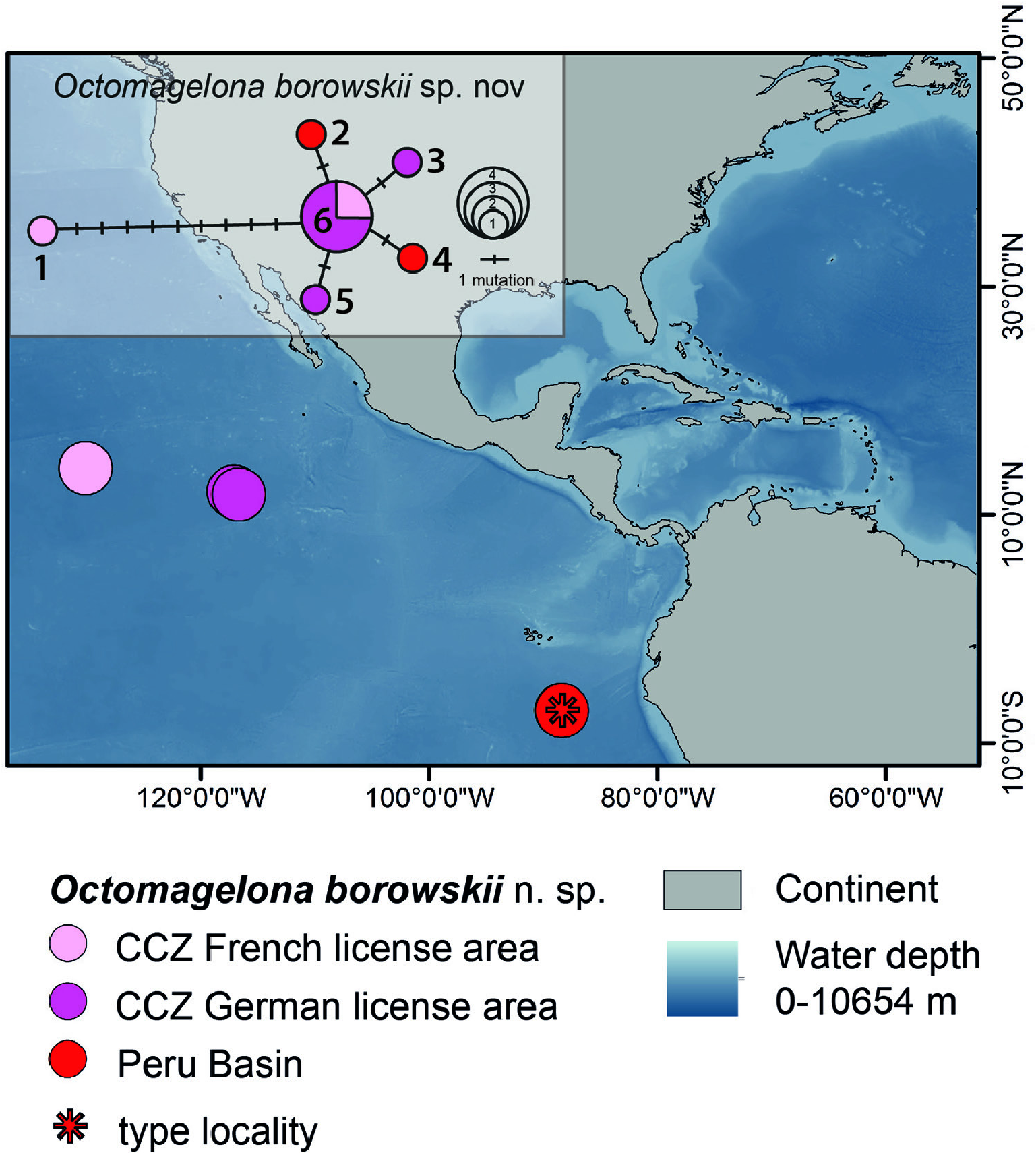
Additional material examined: Central Eastern Pacific, CCZ French licence area, BioNod12, station 101-1, 5055 m (SMF 30531); Central Eastern Pacific, CCZ German license area, BioNod12, station 6-1, 4259 m (SMF 30531, SMF 30532, SMF 30533, SMF 32223, SMF 32224). SE Pacific, Peru Basin (DISCOL area), SO 242-1 (JPIO-DISCOL 1), station 37-1, 4161 m (SMF 30510); station 126-9, 4257 m (SMF 30509). SO 61 (DISCOL 1), station 1276-1, 4153 m (SMF 30511). SO 64 (DISCOL 2), station 1377-1, 4124 m (SMF 30512); station 1403-1, 4166 m (SMF 30514); station 1414-1, 4149 m (SMF 30515); station 1416-1, 4142 m (SMF 30516); station 1419-1, 4156 m (SMF 30517); station 1423-1, 4145 m (SMF 30513). SO 77 (DISCOL 3), station 1449-1, 4148 m (SMF 30524); station 1465-1, 4161 m (SMF 30525); station 1471-1, 4152 m (SMF 30526); station 1476-1, 4174 m (SMF 30523); station 1485-1, 4167 m (SMF 30527); station 1485-1, 4167 m (SMF 32261). – for details and additional specimens see the Supporting Information, Table S2 View Table 2 .
Description: All specimens incomplete, i.e. anterior fragments with only few abdominal segments. Tentacles lost in all specimens, free tentacle fragments only found with two specimens from Peru Basin (SMF 30515, SMF 30521) with four rows of pinnules along tentacle, pinnule length twice diameter of tentacle resulting in frilly appearance. Distal part of tentacle with densely arranged pinnules and free tip.
Prostomium wider than long, length/width/ratio: 0.75 mm / 1.25 mm /0.6 (paratype SMF 30530 View Materials : 0.5 mm / 0.8 mm /0.625, paratype SMF 30528 View Materials : 0.25 mm / 0.55 mm /0.45), truncate, with small frontal horns with free tips, anterior margin straight, smooth. Posterior ends of prostomium bent down forming postero-lateral flaps. Fixed specimens with prostomium pushed up and backward by partly everted bulbous burrowing organ. Eyes absent. Palps lost. First segment (peristomium) achaetous, best visible ventrally ( Fig. 12A, B View Figure 12 ).
Thoracic region with eight chaetigers ( Fig. 12A–F View Figure 12 ). Length 2.19 mm, width decreasing from 0.94 mm at chaetiger 1 to 0.57 mm at chaetiger 8 (paratype SMF 30530 View Materials : length 1.2 mm, width 0.6 to 0.42 mm; paratype SMF 30528 View Materials : length not measured, width 0.45 mm to 0.20 mm). All thoracic parapodia without superior notopodial process [DML of Jones (1963)]. Notopodial lateral lamellae postchaetal, present in chaetigers 1–4 (1–4 in paratypes SMF 30528 View Materials and 30530) filiform with slightly broadened base; decreasing in length from chaetigers 1–4 to less than half length, and absent posteriorly ( Fig. 12A–C View Figure 12 ). In chaetigers 5–8 shallow ridge encircling chaetal fascicles like cuffs ( Fig. 12D, E View Figure 12 ). Neuropodial lateral lamellae postchaetal, present in chaetigers 1–3(4) similar in size and form to notopodial lateral lamellae (slightly smaller in paratype SMF 30528 View Materials ). Gradually decreasing in length from chaetigers 1–4, lacking in posterior segments. Noto- and neuropodia of chaetigers 5–8 forming shallow ridges encircling chaetal fascicles like cuffs. Neuropodia similar to notopodia but slightly larger ( Fig. 12D, E View Figure 12 ). Ventral neuropodial process present in chaetiger 1 only, very small (absent in paratype SMF 30528 View Materials , Fig. 12A–F View Figure 12 ). Thoracic chaetae all long uni- or irregularly bilimbate capillaries, similar in length; most numerous in chaetigers 1–3 (paratype SMF 30528 View Materials with 10–14 in each fascicle), reduced in number posteriorly (?broken) (paratype SMF 30528 View Materials ca. 3–5 in chaetigers 7 and 8). Constriction between chaetiger 8 and 9, i.e. thorax and abdomen ( Fig. 12A, F View Figure 12 ).
Abdomen starting with chaetiger 9. Width 0.63 mm (paratype SMF 30530 View Materials : 0.6 mm; paratype SMF 30528 View Materials : 0.3 mm). Abdominal parapodia without superior notopodial (DML) and ventral neuropodial process (VML) at upper- and lowermost ends of chaetal rows. Noto- and neuropodial lateral lamellae short filiform with slightly broadened base, present in chaetigers 9–11 positioned symmetrically, i.e. sub- and suprachaetal, respectively, at lateral ends of rows of abdominal hooks; increasing in length from chaetigers 9–11 (abdomen broken) ( Fig. 12F View Figure 12 ). Abdominal chaetae tridentate hooded hooks with main fang surmounted by two small teeth; hooks arranged in single line on low parapodial ridge, 6–10 hooks per parapodial ramus, all same size, with three to five each in vis-à-vis arrangement, i. e. noto- and neuropodia with two groups each, teeth facing ( Fig. 12I View Figure 12 , I’). Lateral pouches not observed. Pygidium unknown.
Thorax dorsolaterally with fields of whitish spots, best developed in chaetigers 2–3, less in chaetigers 4–6, situated more laterally; only laterally in chaetiger 7 and 8, mid-dorsum always without spots. Fields of white spots almost touching midventrally in chaetigers 1–3; only few spots in chaetiger 4 and following. Pigment spots in abdominal chaetigers not observed.
Remarks: Until now, Octomagelona bizkaiensis has been known as the single species in the genus originating from about 1000 m depth in the Cap Breton Canyon (NE Atlantic, Gulf of Biscay) (Aguirrezabalaga et al. 2001). The specimens reported here represent the second species for the genus likewise collected from abyssal depths. Additional yet undescribed species have been reported by Mortimer (2019).
Specimens collected in the Pacific from the Peru Basin and the Clarion– Clipperton Fracture Zone do not differ significantly with regard to COI sequences ( Fig. 4 View Figure 4 ). Unfortunately, sequencing of specimens collected from the South Atlantic was unsuccessful and we also do not have any genetic information for O. bizkaiensis from the Bay of Biscay.
Octomagelona borowskii sp. nov. differs from O. bizkaiensis by slightly more distinct prostomial horns, a thorax decreasing significantly in width posteriorly, the presence of filiform postchaetal noto- and neuropodial lateral lamellae only in thoracic chaetigers 1–4 replaced by parapodial cuffs encircling chaetal fascicles in more posterior thoracic chaetigers, while parapodia in O. bizkaiensis appear completely reduced. Pigmentation is less dense and less pronounced in O. borowskii sp. nov. and spots are whitish instead of dark pigment granules as in O. bizkaiensis .
One specimen collected from the CCZ ( SMF 30533 View Materials ) showed pigmentation spots in dense patches dorsolaterally on chaetigers 2–7, and on chaetigers 2–5 also laterally; ventrally pigmentation spots are scaưered and more dense in anterior thorax .
Etymology: The specific name refers to Dr. Christian Borowski, who studied the polychaetes collected during the initial phase of project DISCOL for his Ph.D. project. He was the very first to observe magelonids with only eight thoracic chaetigers and described them tentatively as ‘ Magelona sp. A’ (Borowski 1996).
Distribution: Central Pacific (CCZ, 4128–5055 m), SE Pacific ( Peru Basin, 4124–4257 m).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.