
Spiophanes cf. longisetus Meissner, 2005
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlad069 |
|
publication LSID |
lsid:zoobank.org:pub:65B60DD3-64C9-4262-B7B2-74DA4D3D889F |
|
DOI |
https://doi.org/10.5281/zenodo.10497570 |
|
persistent identifier |
https://treatment.plazi.org/id/743EE917-FFE8-FFAE-FF05-24FB1895FCF6 |
|
treatment provided by |
Plazi |
|
scientific name |
Spiophanes cf. longisetus Meissner, 2005 |
| status |
|
Spiophanes cf. longisetus Meissner, 2005 View in CoL
( Fig. 18 View Figure 18 )
Spiophanes longisetus Meissner, 2005: 45–48 View in CoL , figs 26–28, table 1.
Spiophanes kroyeri Grube, 1860 View in CoL . – Hartman and Fauchald, 1971: 105–106.
Material examined: Central Atlantic Ocean, Mid-Atlantic Ridge, East, SO 237 (VEMA-Transit), stn 2-6, EBS, 12 Jan 2015, 5520 m, one af, tissue sample ( ZMH-P 28149 ), one af, tissue sample ( ZMH P-28150); stn 2-7, EBS, 20 Dec 2014 , 5507 m, one af ( ZMH P-28154), one af ( ZMH-P28155 ), one af ( SMF 30642 View Materials , SEM 1330 ), one af ( SMF 30643 View Materials , SEM 1331 ), one mf ( SMF 30644 View Materials ); stn 4-8, EBS, 26 Dec 2014 , 5725 m, one af, tissue ( ZMH P-28156); stn 4-9, EBS, 27 Dec 2014 , 5733 m, two af ( ZMH P-28157); stn 6-7, EBS, 2 Jan 2015 , 5079 m, four af ( SMF 30641 View Materials , SEM 1342 ); stn 6-8, EBS, 2 Jan 2015 , 5079 m, one af, tissue ( ZMH-P 28160 ). Mid-Atlantic Ridge, Central, SO 237 (VEMA-Transit), stn 8-4, EBS, 6 Jan 2015 , 5176 m, one af, tissue ( SMF 30640 View Materials , SEM 1327 ). Mid-Atlantic Ridge, West, SO 237 (VEMA-Transit), stn 9-8, EBS, 12 Jan 2015 , 5004 m, one af, tissue ( ZMH-P 28162 , SEM), one af, tissue ( SMF 30639 View Materials ), one af, tissue ( ZMH P-28164); stn 11-4, EBS, 14 Jan 2015 , 5108 m, one af ( SMF 30638 View Materials ). SW Atlantic Ocean, Brazil Basin N, M 79-1 ( DIVA 3 ), stn 604-1, EBS, 31 Aug 2012 , 5180 m, one af, tissue ( SMF 30645 View Materials ), one af, tissue ( SMF 30646 View Materials ). – for details and additional specimens see the Supporting Information , Table S2 View Table 2 .
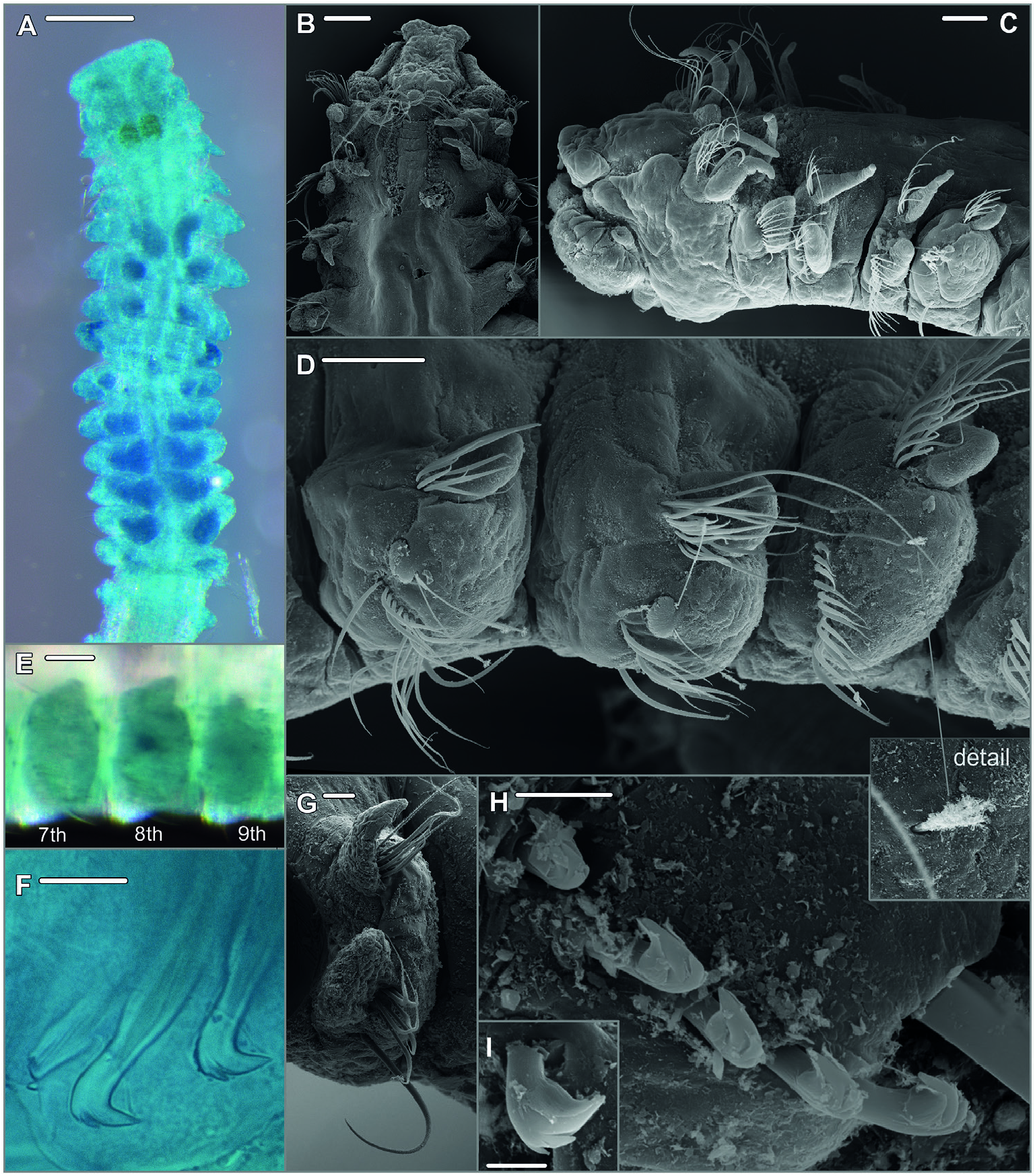
Description: (Focusing on most important characters for specimens examined in the course of the present study.) Specimens all anterior fragments with up to 23 chaetigers, between 0.2 and 0.9 mm in width, and up to 6.4 mm long. Prostomium bell-shaped with straight anterior margin extending into short anterolateral projections ( Fig. 18A, B View Figure 18 ), short, stout cirriform occipital antenna with rounded tip ( Fig. 18B, C View Figure 18 ). Dorsal ciliated organs as dorsal ciliated grooves posterior to the prostomium (ciliation detectable with SEM), if viewed with LM appearing as thick, straight, double lines of ochre or yellowish colour reaching the end of the 2nd chaetiger ( Fig. 18A, B View Figure 18 ), sometimes outer margins of ciliated grooves demarcated and nuchal organ then appearing as pair of short U-shaped double lines. Ventral sabre chaetae from chaetiger 4, oħen of imposing length ( Fig. 18C, D, G View Figure 18 ). Chaetal spreader of the ‘0 + 1 type’ with semicircular glandular opening developed in chaetigers 5–7, in chaetiger 8 chaetal spreader present but with only small hole-like opening ( Fig. 18D View Figure 18 ); glandular organ of chaetigers 9–14 opens as a lateral vertical slit, without chaetal spreader. Bacillary chaetae as thin hirsute bristles can be exposed on chaetigers 5–8 though small opening of glandular organ on chaetiger 8 allowing only the protrusion of distal tips of very few bacillary chaetae ( Fig. 18D View Figure 18 ). Few single ciliated patches randomly present in parapodia of the middle body region or completely absent. Ventrolateral intersegmental genital pouches absent. Posterior region starting at chaetiger 15 with first presence of neuropodial quadridentate hooks with main fang surmounted by single tooth and two uppermost smaller teeth in parallel position, hooks with half-hood from the tip of the main fang to the shaħ, usually four to five hooks, in some juveniles only three hooks arranged in one row ( Fig. 18F, H, I View Figure 18 ); single, thin accompanying capillaries oħen present, observed in position next to sabre chaeta; notopodia with slightly granulated capillaries arranged in a tuħ, among those few strikingly long capillaries. Pygidium not observed in examined specimens (all anterior fragments).
Pigmentation: All examined specimens pale without pigmentation, only nuchal organ with yellowish or ochre pigment as in related species ( Fig. 18A View Figure 18 ). According to original description for S. longisetus Meissner, 2005 yellow to orange pigment in neuropodia of chaetigers 11–14 present. Remnants of faint brownish pigment in neuropodia 11–14 in few specimens discernible (e.g. SMF 30645), but usually absent.
Methyl green staining pattern: Chaetigers of the anterior middle body region, and especially their subepidermal glandular organs, most intensely stained and also most persistently stained compared to other parts of the body. In chaetiger 8, lateral part of the neuropodium most intensively stained, in lateral view observed as dark circular area ( Fig. 18A, E View Figure 18 ).
Biology: Information about reproduction and development not available, because none of the studied specimens was bearing gametes.
Remarks: The species is morphological very similar to other congeners from the deep-sea discussed in this paper: S. australis sp. nov. (found in the SW Atlantic and adjacent Antarctic waters) and S. pacificus sp. nov (collected from the Pacific Ocean). All three species are morphologically best distinguished by the distribution of lateral neuropodial ciliated patches along the middle body region, which are arranged in distinct paưerns in S. australis sp. nov. and S. pacificus sp. nov. but are only randomly found as single patches in S. cf. longisetus , or also completely absent in the laưer. The number of neuropodial hooks in posterior chaetigers is with (3)4–5 highest in S. cf. longisetus whereas in S. australis sp. nov. (3–)4 hooks are present and in S. pacificus sp. nov. usually not more than three hooks are found. The species can also be distinguished based on information from molecular markers ( COI). However, the identity of specimens here referred to as S. cf. longisetus is not entirely resolved. The problem is mainly caused by the lack of information on molecular markers for S. longisetus Meissner, 2005 from type material or other specimens from the type locality. Moreover, our search in public sources (GenBank, BOLD) for sequences in good agreement with our putative species here referred to as S. cf. longisetus was unsuccessful. Based on what we know today molecular information is indispensable for solving the problem of the species identity since another Spiophanes species from the abyssal NE Atlantic is known: S. abyssalis Maciolek, 2000 . Records for this species come from two different localities in the Bay of Biscay and the type locality close to the Canary Islands. Spiophanes abyssalis is morphologically extremely close to S. longisetus . Morphological differences concern the hood of the neuropodial hooks which are described as rudimentary and hard to observe in S. longisetus whereas they are clearly visible in S. abyssalis ( Meissner 2005) . In the laưer species, four to five hooks were observed, in S. longisetus three to five. Spiophanes longisetus can also be identified by the presence of long granulated notopodial chaetae from chaetiger 14 whereas for S. abyssalis posterior notopodial chaetae are described as simple narrowly sheathed capillaries [updated species description in Meissner (2005)]. Also, in the original description Maciolek (2000) describes the notochaetae as comparatively short and moreover states the presence of two eyes in the holotype. Since the here examined specimens from the central Atlantic present very long notopodial capillaries, up to five neuropodial hooks and no eyes, we here refer to them as S. cf. longisetus . However, a more reliable conclusion will be possible if additional molecular information becomes available and a subsequent review of the morphology of all involved species, especially in regard to the newly discovered ciliated patches and the number of neuropodial hooks in relation to specimen size, can be undertaken.
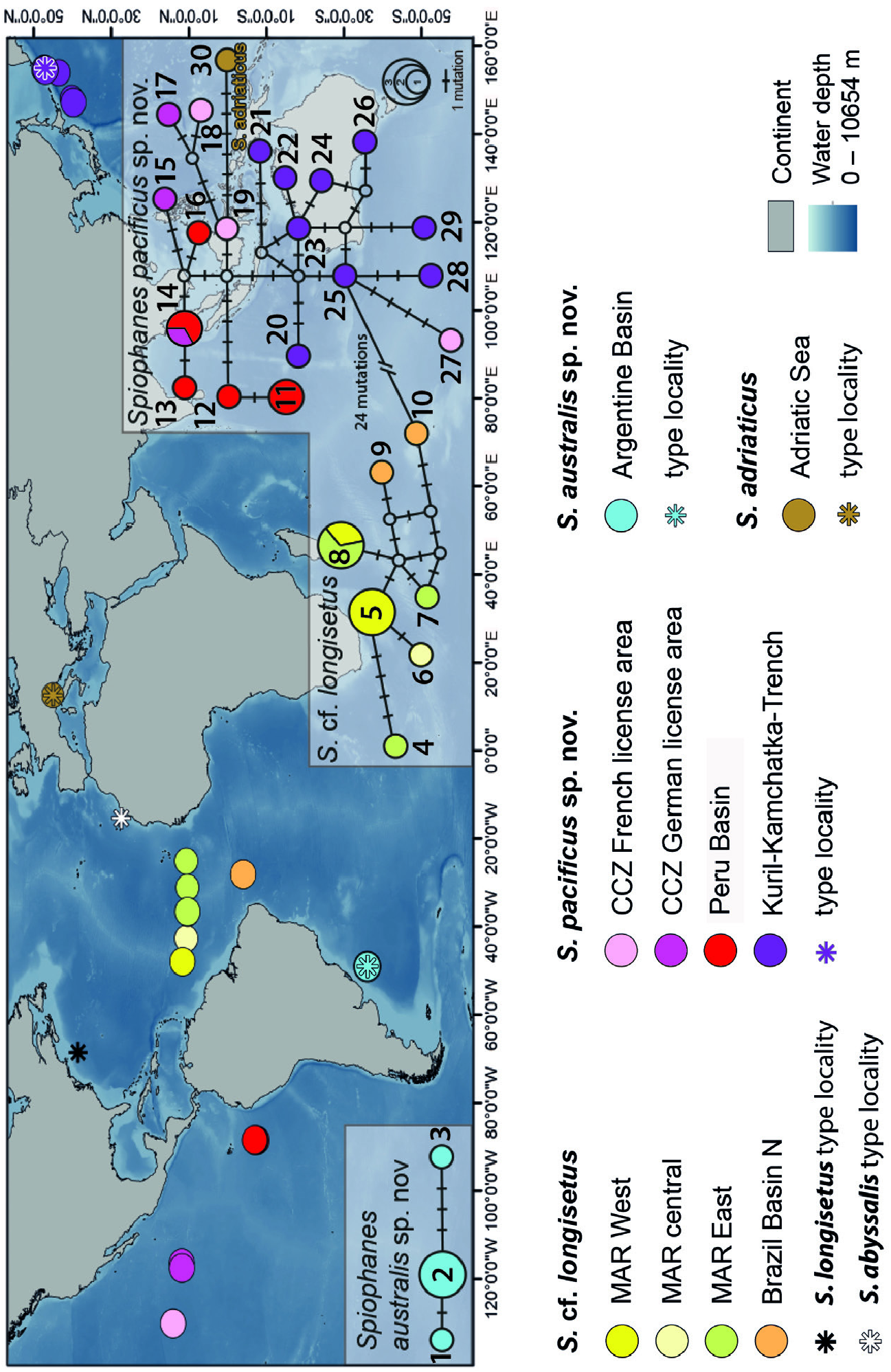
Distribution: Spiophanes longisetus Meissner, 2005 has been described from localities in the NW Atlantic Ocean ( Meissner 2005). Type material and additional non-type material studied while describing the species all came from slope and abyssal depths off New England and Bermuda in the NW Atlantic Ocean ( Meissner 2005). Specimens studied in the course of the present study and tentatively suggested to belong to S. longisetus came from the VEMA fracture zone of the Mid-Atlantic Ridge and abyssal plains east and west of it, as well as from the Brazil Basin in the northern South Atlantic Ocean ( Fig. 5 View Figure 5 ). Water depths were 3753–4663 m in the Western North Atlantic, 5004–5733 m at locations near the Mid-Atlantic Ridge, and 5200 m in the Brazil Basin. As soon as new information on genetic markers for specimens from the type locality becomes available and the uncertainty concerning the identity of the different specimens can be dispelled, the distribution of S. longisetus has to be revalidated.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Spiophanes cf. longisetus Meissner, 2005
| Meissner, Karin, Schwentner, Martin, Göưing, Miriam & Fiege, Thomas Knebelsberger and Dieter 2023 |
Spiophanes longisetus
| Meissner K 2005: 48 |