
Ephesiella australiensis Hartmann-Schröder, 1982
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4000.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:7EDEDAEE-642C-4F9D-A04D-141815D73343 |
|
DOI |
https://doi.org/10.5281/zenodo.5667445 |
|
persistent identifier |
https://treatment.plazi.org/id/81007D79-8D49-256F-FF0F-FD7FFCAE1E6E |
|
treatment provided by |
Plazi |
|
scientific name |
Ephesiella australiensis Hartmann-Schröder, 1982 |
| status |
|
Ephesiella australiensis Hartmann-Schröder, 1982 View in CoL
Figs 1 View FIGURE 1 A–C, 2, 3, 4A–B, 5A–D
Ephesiella australiensis Hartmann-Schröder, 1982: 80 View in CoL –81, Figs 84–87.
Material examined. Holotype: Cervantes, Western Australia, Australia. ZMH P.16773, sandy beach between Posidonia , 24 Oct 1975. Paratype: (1 spec.) same details. Additional material. Western Australia: AM W.42699 (1 spec.), North West Shelf, 19° 28' S, 118° 55' E, 39 m, 26 Apr 1983; AM W.42700 (1 spec.), 2 km west of Angel Island, Dampier Archipelago, 20° 29' 46" S, 116° 47' 29" E, 10 m, 0 4 Aug 2000, dead coral; AM W.42701 (2 specs, one for SEM), Angel Island, Dampier Archipelago, 20° 27' 41" S, 116° 47' 31" E, 14 m, 0 5 Aug 2000, dead coral; AM W.42702 (2 specs, one for SEM), south west Enderby Island, 20° 37' 18" S, 116° 27' 23" E, 14 m, 0 8 Aug 2000, dead coral; AM W.42703 (2 specs), north west end Legendre Island, 20° 21' 13" S, 116° 50' 26" E, 21 m, 30 Jul 2000, dead coral; AM W.42704 (4 specs), 1 km north east of Delambre Island, Dampier Archipelago, 20° 25' 43" S, 117° 05' 07" E, 14 m, 0 7 Aug 2000; NMV F.162479 (1 spec.), off Pelsart Island, near Geraldton, 29° 00' 10" S, 113° 46' 26" E, 409 m, 0 1 Aug 2005. Northern Territory: NTM W.10209 (1 spec.), Darwin Harbour, 12° 29' 3" S, 130° 50' 10"E, 7m, 1993; NTM W.15191 (1 spec.), Darwin Harbour, 12° 23' 15" S, 130° 49 32' 6" E, 1994; NTM W.15197 (1 spec.), Darwin Harbour, 12° 28' 10" S, 130 46 '15" E, 5 m, 1994; NTM W.20663 (1 spec. for SEM), Arafura Sea, 18° 22' 30" S, 133° 41' 48" E, 187 m, 2005; NTM W.24312 (1 spec.), Joseph Bonaparte Gulf, NT, Australia, 12° 20' 56" S, 129° 58' 40" E, 44 m, 2009; Queensland: AM W.202140 (2 specs), near mouth of Althaus Creek, Halifax Bay, north of Townsville, 19° 10' S, 146° 37' E, Jul 1977; AM W.42693 (1 spec. for SEM); New South Wales: AM W.28133 (1 spec.), east of Wollongong, 34° 28' S, 151° 02' E, 100 m, 28 Mar 1994, in Globigerina ; AM W.42688 (1 spec. for SEM), Cape Three Points, south-east of Third Point, 33° 31' 55" S, 151° 24' 58" E, 30 m, 0 6 May 2007, sand from around large boulders; AM W.42711 (1 spec.), Bass Point, 34° 36' S, 150° 54' E, 65 m, 25 Jun 1990; AM W.42712 (1 spec.), same details; AM W.42728 (3 specs), east of Malabar, 33° 58' 41" S, 151° 18' E, 81.4 m, 23 Aug 1995. Victoria: NMV F.132625 (1 spec.), 43 km SE of Port Albert, Eastern Bass Strait, 38° 53' 42" S, 147° 06' 30" E, 58 m, 18 Nov 1981; NMV F.132637 (1 spec.), 63 km E of North Point, Flinders Island Eastern Bass Strait, 39° 44' 48" S, 148° 40' 36" E, 124 m, 14 Nov 1981; NMV F.132642 (1 spec. for SEM), 50 km SW of Warrnambool, Western Bass Strait, 38° 49' 30"S, 142° 35' 24"E, 89 m, 21 Nov 1981.
Diagnosis. Microtubercles absent or present on first chaetigers. Palps and lateral antennae digitiform, median antenna spherical. Tentacular cirri ellipsoid or inconspicuous. Eyes not observed. Parapodia with 4–6 parapodial papillae; compound or semi-compound chaetae with blades 1.5–2.5 times as long as maximum width on mid-body chaetigers; hooks absent or present on first chaetiger.
Re-description.
Measurements and general morphology. Holotype 1.4 mm long, 0.4 mm wide, with 18 chaetigers. Body elongated, sub-quadrangular in section, with slightly convex dorsum. Anterior end bluntly rounded, slightly narrowing along posterior segments. Segmentation inconspicuous, tegument with transverse wrinkles. Preserved specimen lacking pigmentation.
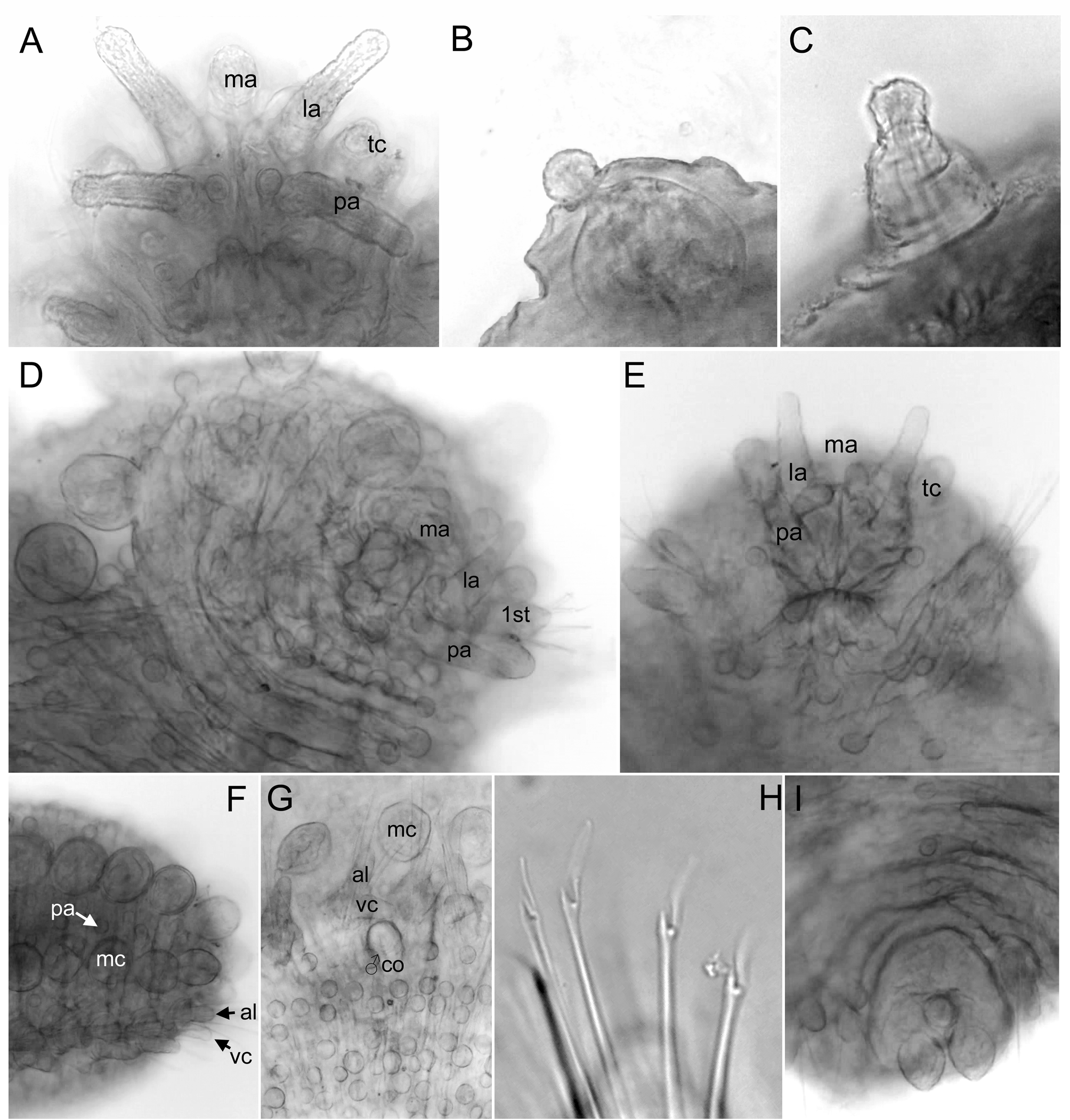
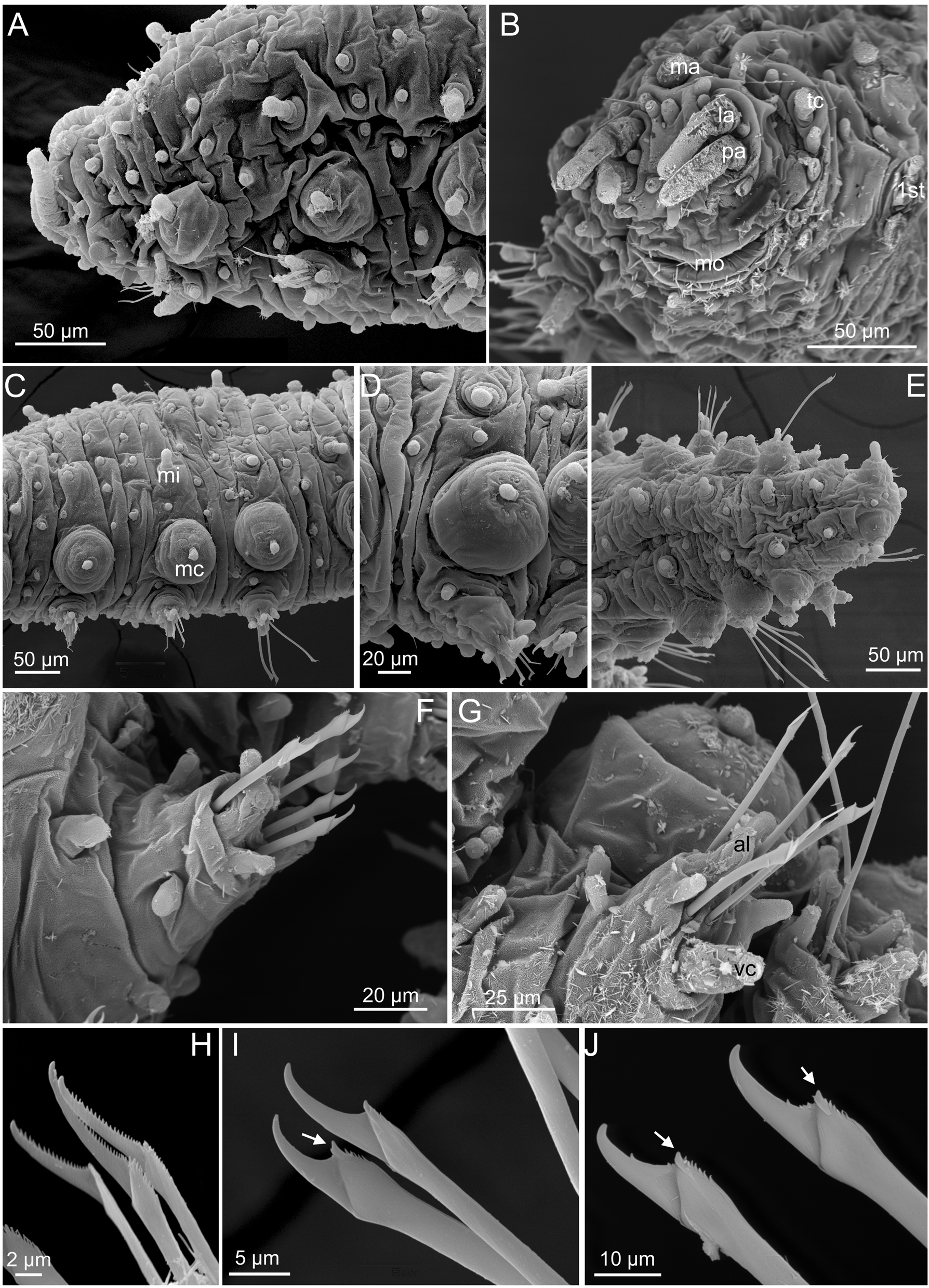
Head. Prostomium with five short appendages, including a pair of digitiform palps in ventral-most position, a pair of lateral antennae, similar in shape and size to palps, and a median antenna, shorter (one third) than lateral antennae and resembling a large semi-spherical papilla ( Figs 1 View FIGURE 1 A–B, 2A–B, 3A, G). Two rounded small papillae between palps, two between median and lateral antennae and two between lateral antennae and mouth ( Figs 1 View FIGURE 1 A, 2B). A pair of tentacular cirri similar in shape and size to lateral antennae and palps and several scattered papillae similar to prostomial ( Fig. 2 View FIGURE 2 B).
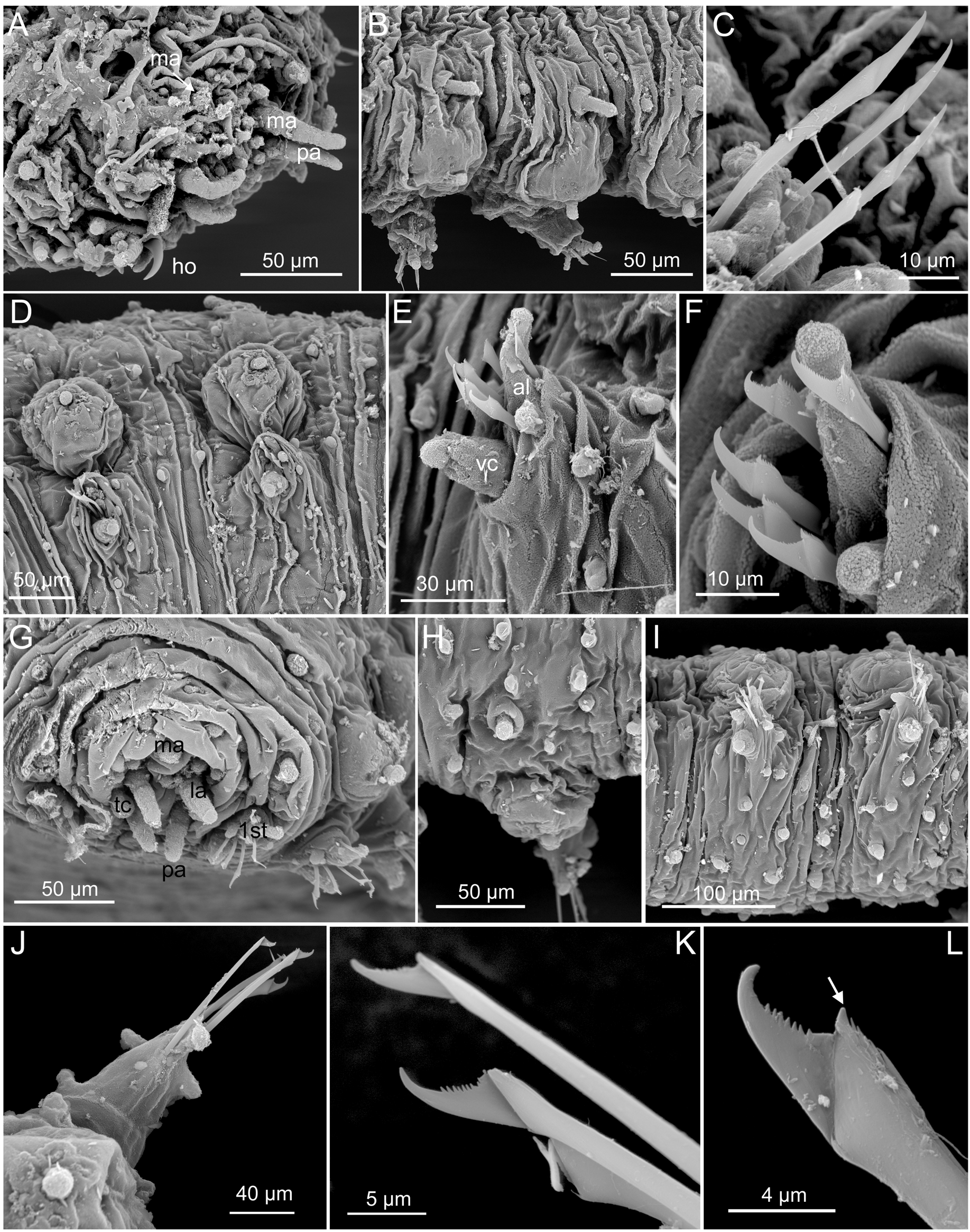
Tubercles. First chaetiger with two dorsal macrotubercles; microtubercles absent ( Figs 2 View FIGURE 2 A, 3G). Following chaetigers each with two dorsal macrotubercles arranged in two dorso-lateral longitudinal rows, and two microtubercles forming two longitudinal rows between the macrotubercles ( Figs 2 View FIGURE 2 C, 4A). Macrotubercles sessile and spherical, each provided with a digitiform terminal papilla ( Figs 1 View FIGURE 1 A, 2A, C, D, 3B, D); with groups of pores around terminal papilla ( Fig. 2 View FIGURE 2 D). All macrotubercles similar in shape and size, slightly increasing in size to chaetiger 4 and decreasing in posterior chaetigers. Microtubercles with digitiform terminal papillae shorter ( Fig. 1 View FIGURE 1 C), or longer ( Figs 2 View FIGURE 2 A, C, E, 3B, H) than collar. Spherical papillae over dorsum, arranged in three transversal rows per chaetiger, with around 15–20 papillae on each mid-body chaetigers, including 2–3 papillae between macrotubercles and parapodia ( Figs 2 View FIGURE 2 C, 4A). Ventral surface with spherical papillae, arranged in four more or less regular transversal rows, with a total of 20–25 per segment, in mid-body; numbers decreasing towards posterior end ( Figs 3 View FIGURE 3 I, 4B). Body epithelium with ellipsoid granules (e.g. Fig. 3 View FIGURE 3 F).
Parapodia. Parapodia sub-conical, increasing in size towards chaetiger 3, around 1–2 times longer than wide; acicular lobe projecting distally anterior to chaetae, resembling other parapodial papillae or slightly longer, ventral cirri bottle-shaped projecting as long as acicular lobe ( Figs 2 View FIGURE 2 A, C–D, F–G, 3B, E–F, I–J). Anterior parapodia with four hemispherical papillae: one anterior, one anterio-ventral, one anterior dorsal and one posterior, in addition to the acicular lobe. Mid-body parapodia with six semi-spherical papillae, all similar in size, in addition to the acicular lobe: one anterior dorsal, one anterior, near the acicular lobe, one anterior ventral, two posterio-dorsal (one of them distal) ( Figs 2 View FIGURE 2 F–G, 3E, J, 5A–D).
Chaetae. First chaetiger apparently with hooks absent. Compound or semi-compound chaetae in all chaetigers, arranged in a curved transverse row around acicular lobe and numbering 4–6 per fascicle ( Figs 2 View FIGURE 2 F, G, 3E, F). First and second chaetigers with well serrated long blades, 5–6 times longer than wide ( Fig. 2 View FIGURE 2 H). Chaetae from chaetiger 3 with shafts widened distally, a larger distal tooth and fine spinulation; blades twice as long as wide, recurved and with smooth or few serration of the inner edges, with dorso-ventral gradation in the width and length ( Fig. 2 View FIGURE 2 I–J, 3C, K–L).
Pygidium . Pygidium terminal, with mid-ventral digitiform anal cirrus and a pair of dorsal anal cirri, similar in shape but slightly smaller than macrotubercles ( Fig. 2 View FIGURE 2 E).
Internal features. Eyes or muscular pharynx not observed.
Reproductive features. Gametes or ‘copulatory organs’ not observed on either type or additional material. Some gravid females (e.g. NMV F.132637) are completely filled with discoid eggs, 250 µm of diameter.
Variation. The holo- and the paratype are the smallest individuals reported. Other preserved specimens range between 2.2 and 22 mm long, 0.2 and 1.8 mm wide and had between 30 and 64 chaetigers. The relative length of the prostomial antennae is constant among specimens, palps and lateral antennae are similar in shape and size, and larger than median antenna. Some specimens have small or even inconspicuous tentacular cirri while these are digitiform and clearly visible in others. Differences in the absence/presence of microtubercles on the first chaetiger have been observed even within type material (the paratype bears microtubercles on first chaetiger). Chaetae also showed variation among the material examined. Some specimens from Northern Territory bear hooks on first chaetiger ( Fig. 3 View FIGURE 3 A) and most specimens, regardless their origins, bear semi-compound chaetae with joined shaft and blade although recognisable (e.g. Fig. 3 View FIGURE 3 C, F, L). Diversity in the relative length/width of the blades was also observed within ( Fig. 2 View FIGURE 2 H–J) and among specimens (being the Northern Territory specimens the one presenting longer blades, Fig. 3 View FIGURE 3 C). Moreover, a few specimens collected from Victoria, show enlarged macrotubercles with embedded microtubercles and some papillae, what is interpreted as an artefact due to swollen macrotubercles ( Fig. 5 View FIGURE 5 D). The number (4–6) and arrangement of parapodial papillae show little variation among specimens ( Fig. 5 View FIGURE 5 A– D).
Remarks. This species was originally described based on the combination of the following features: absence of eyes, absence of hooks on first chaetiger, median antenna shorter than lateral, 4–5 parapodial papillae and compound chaetae with blades 1.5–2 times as long as maximum width ( Hartmann-Schröder 1981). The specimens collected along the Australian coast mostly fit on this description but present slight variation on other morphological traits, as indicated above, not congruent with particular geographical patterns. It is therefore difficult, at this stage, to determine if there are more than one Ephesiella species present in Australia, despite the large geographical range and environmental diversity reported here for the species. The genus Ephesiella is a homogenous group of sphaerodorids currently with 15 species described and reported from around the world. The species boundaries and their diagnostic features need reassessment considering intraspecific variability which is not often taken into account ( Desbruyères 1980; Hartmann-Schröder 1981). For the time being, all the specimens collected in Australia have been considered as a single entity but this hypothesis should be tested, desirably with molecular data.
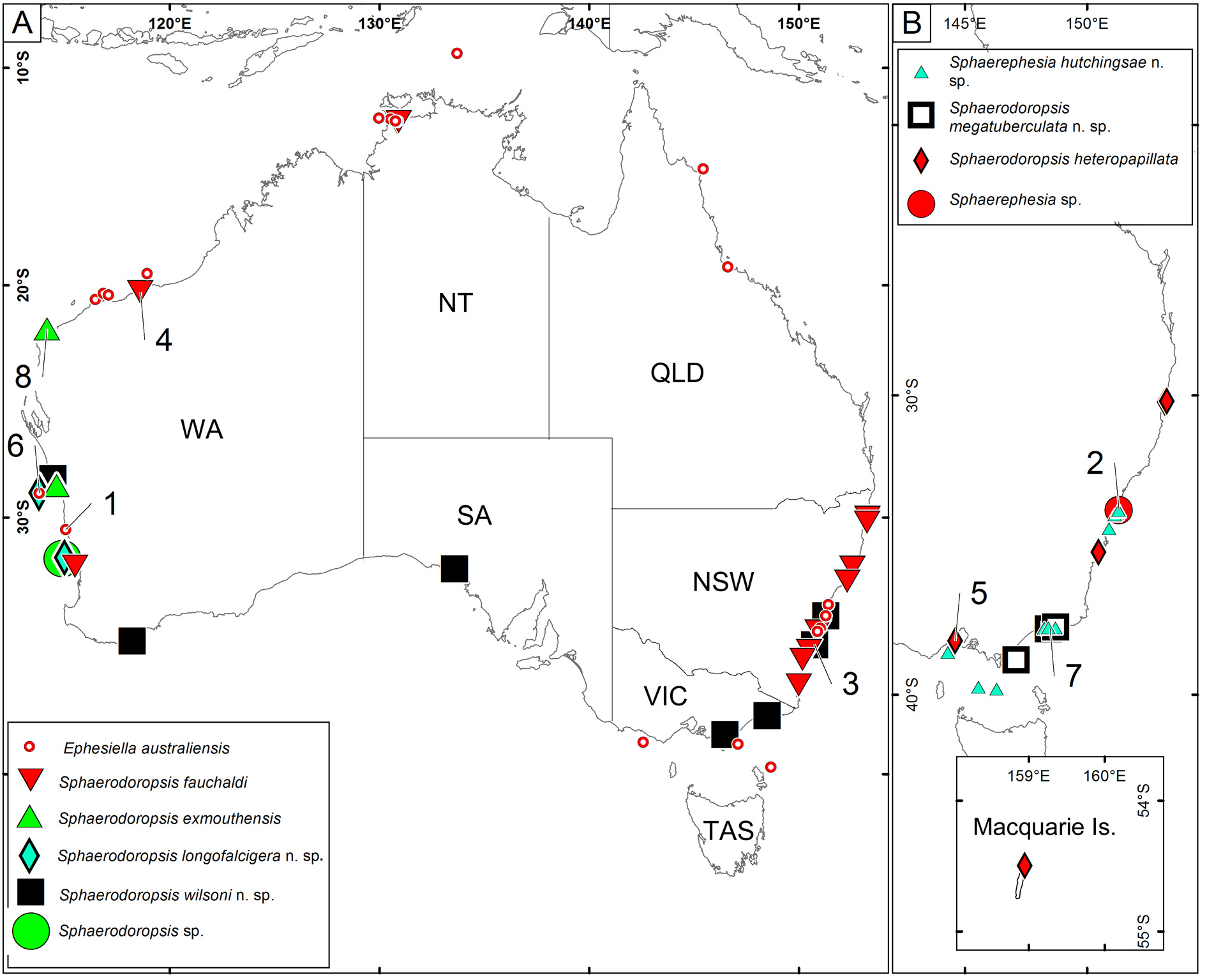
Type locality. Cervantes, Western Australia ( Fig. 15 View FIGURE 15 ).
Distribution. Western Australia, Northern Territory, Queensland, New South Wales and Victoria (Hartmann- Schröder 1981; Wilson & Bakken 2003; Capa & Rouse 2015; Fig. 15 View FIGURE 15 ).
Habitat. Sand, sea-grass beds, dead coral, 10–400 m depth.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ephesiella australiensis Hartmann-Schröder, 1982
| Capa, Maria & Bakken, Torkild 2015 |
Ephesiella australiensis Hartmann-Schröder, 1982 : 80
| Hartmann-Schroder 1982: 80 |