
Arachnothelphusa merarapensis, Grinang & Min & Ng, 2015
|
publication ID |
https://doi.org/ 10.5281/zenodo.4502502 |
|
publication LSID |
lsid:zoobank.org:pub:4CC4DC84-8F6E-4524-9D3F-BAA3C2DAF588 |
|
DOI |
https://doi.org/10.5281/zenodo.4776189 |
|
persistent identifier |
https://treatment.plazi.org/id/1822C659-42CD-47F7-911F-D4A469B40154 |
|
taxon LSID |
lsid:zoobank.org:act:1822C659-42CD-47F7-911F-D4A469B40154 |
|
treatment provided by |
Carolina |
|
scientific name |
Arachnothelphusa merarapensis |
| status |
sp. nov. |
Arachnothelphusa merarapensis View in CoL , new species
( Figs. 1–3 View Fig View Fig View Fig )
Material examined. Holotype: male (22.5 × 16.8 mm) ( ZRC), water-filled tree-hole, ca. 100 cm above ground, steep dipterocarp forest, Merarap Hot Spring Resort , Lawas, northern Sarawak, Malaysia, Borneo, 4 o 22’25.4”N 115 o 26’10.1”E, 485 m asl, coll. J. Grinang & Y.M. Pui, 31 October 2014 GoogleMaps . Paratype: 1 female (19.9 × 15.2 mm) ( SBC.C.00376), water-filled hole of tree buttress, ca. 90 cm above ground, steep dipterocarp forest, Merarap Hot Spring Resort , Lawas, northern Sarawak, Malaysia, Borneo, 4 o 22’16.5”N 115 o 26’12.4”E, 494 m asl, coll. J. Grinang & Y.M. Pui, 1 November 2014 GoogleMaps ; 1 female (22.3 × 16.1 mm) ( SBC.C.00377), water-filled hole of tree buttress, ca. 30 cm above ground, same data as paratype, coll. Y.M. Pui, 27 February 2013 GoogleMaps .
1 Institute of Biodiversity and Environmental Conservation, Universiti Malaysia Sarawak, 94300 Kota Samarahan, Sarawak, Malaysia; Email:gjongkar@ibec.unimas. my (* corresponding author)
2 Lee Kong Chian Natural History,Faculty of Science,National University of Singapore, 6 Science Drive 2, 117543 Singapore.
© National University of Singapore
ISSN 2345-7600 (electronic) | ISSN 0217-2445 (print)
Comparative material. Arachnothelphusa kadamaiana ( Borradaile, 1900) : Holotype female (18.9 × 13.5 mm) ( SMF 4281 About SMF ), Kadamian River , Sabah, Malaysia, Borneo ; 1 male (20.1 × 14.9 mm) ( SMF 4282 About SMF ), same data as holotype; 1 female (23.2 × 17.1 mm) ( ZRC 2009.0094 View Materials ), Poring, Basin 1A, Sabah, Malaysia, Borneo, coll. R. F. Inger et al., 12 August 1992.
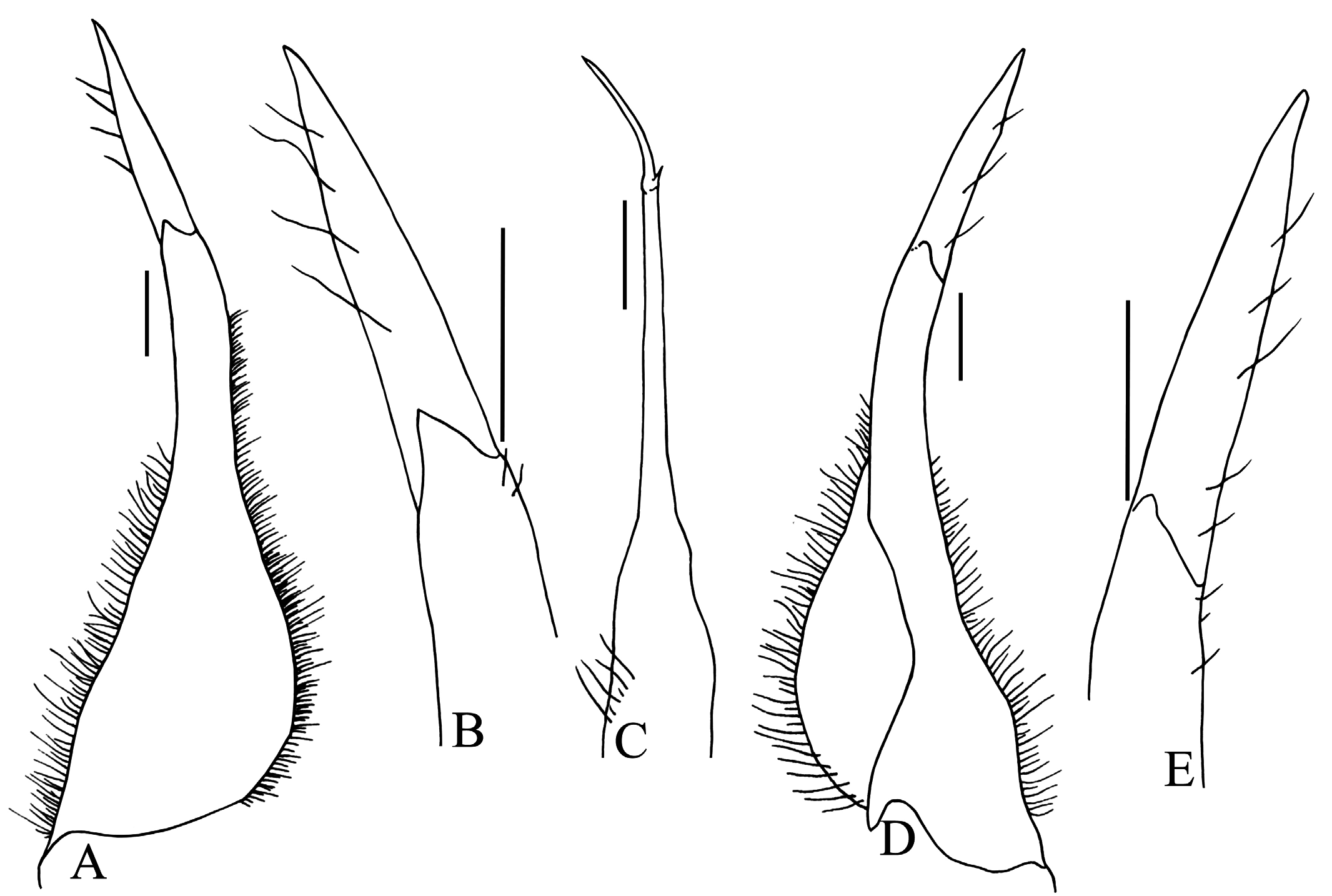
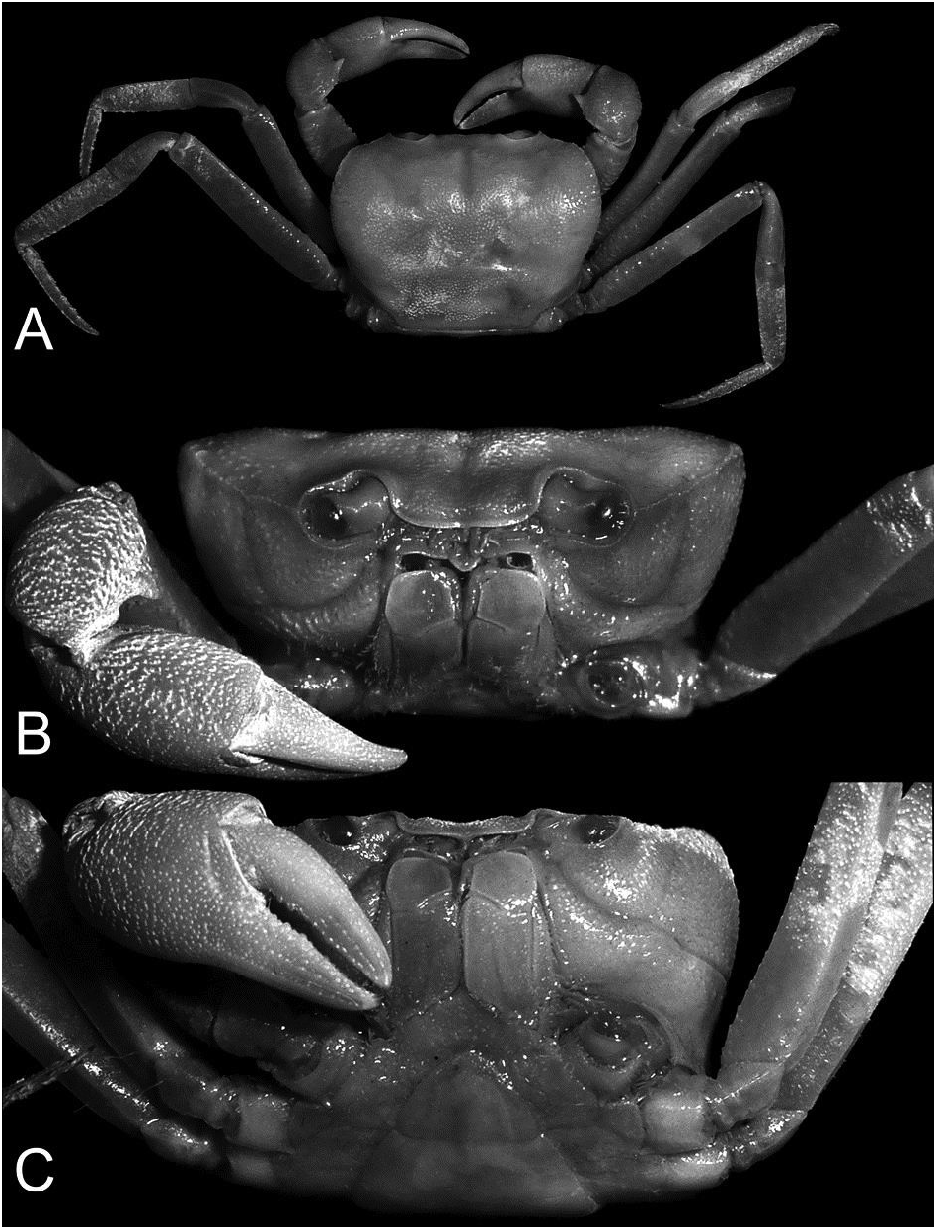
Diagnosis. Carapace surface convex, rugose, finely granular; regions distinct, branchial regions slightly inflated; epigastric and postorbital cristae distinct; cervical and H-shaped gastric grooves deep, not confluent; anterolateral margins convex, serrated; antero- and posterolateral regions prominently rugose, covered with numerous coarse granules; epibranchial tooth acutely triangular, distinctly separated from external orbital angle by wide, deep cleft; external orbital angle acutely triangular, outer margin slightly concave, distinctly serrated ( Figs. 1 View Fig A–C, 3). Ischium of third maxilliped rectangular, oblique median sulcus deep ( Fig. 1F View Fig ). Outer surfaces of chelipeds distinctly rugose; carpus rugose, with granules, inner angle with broadly triangular tooth; cutting edge of pollex with larger median tooth ( Fig. 1A, G, H View Fig ). Ambulatory legs long; merus unarmed, dorsal margin gently serrated; outer surface rugose; fourth ambulatory merus subequal to length of carapace ( Fig. 1A, E View Fig ). Male abdomen T-shaped, lateral margins of somites 5 and 6 distinctly concave; somite 6 subequal to length of telson; lateral margins of telson concave ( Fig. 1D View Fig ). G1 slender, sinuous, gently curving outwards; terminal segment cylindrical, tapering, about half of length of subterminal segment ( Fig. 2A, B, D, E View Fig ). G2 with short distal segment, about a quarter length of basal segment ( Fig. 2C View Fig ).
Size. All species of Arachnothelphusa are relatively small, with the largest recorded male a freshly moulted specimen of A. terrapes measuring 25.7 mm by 18.6 mm ( Ng, 1991). The holotype male of A. merarapensis , new species, is 22.5 mm by 16.8 mm.
Variation. The two female paratypes agree well with the holotype male, differing only in minor non-sexual characters: the cervical and H-grooves are proportionately deeper in the larger female, and the ambulatory legs slightly longer. The antero- and posterolateral regions also have relatively more coarse granules in the larger female specimen.
Etymology. The species is named after Merarap, the type locality where it was collected.
Colour. In life the carapace, chelipeds, and walking legs are bright purple; the ventral surfaces of the ambulatory legs and chelipeds are lighter coloured, and the distal parts of the fingers are cream-white ( Fig. 3A, B View Fig ).
Remarks. The high density of granules and roughness of the dorsal carapace surface of A. merarapensis ( Fig. 1A, B View Fig ) easily distinguishes it from its other congeners which have a relatively smoother dorsal carapace surface covered with small granules and scattered striae (cf. Fig. 5 View Fig ; De Man, 1899: pl. 9 fig. 11; Ng & Goh, 1987: pl. 3A; Ng, 1991: figs. 1, 3–5 [incorrectly labelled as fig. 5 with next two figure captions not printed]). The acutely triangular external orbital tooth separated by a wide and deep cleft is diagnostic for A. merarapensis ( Fig. 1A, B View Fig ). In A. melanippe the epibranchial tooth is distinct but relatively small and separated from the broadly triangular external orbital tooth by a small cleft (cf. De Man, 1899: pl. 9 fig. 11; Ng, 1991: fig. 1); in A. kadamaiana the epibranchial tooth is low and the external orbital tooth is broad, with these two teeth appearing almost confluent with hardly any indication of a cleft ( Figs. 5A View Fig , 6A View Fig ); in A. rhadamanthysi the external orbital and epibranchial teeth are both relatively broad due to the wide carapace ( Ng & Goh, 1987: pl. 3A, B); and in A. terrapes there is a deep and broad U-shaped cleft separating the epibranchial tooth from the obtuse and truncate external orbital angle ( Ng, 1991: fig. 3 [incorrectly printed as fig. 5]). The terminal segment of the G1 is long in A. merarapensis , about half the length of the subterminal segment ( Fig. 2A, B, D, E View Fig ); it is about one-third the length of the subterminal segment in A. melanippe (cf. Ng, 1991: fig. 2C–F), and only a quarter of the length of the subterminal segment in A. kadamaiana and A. terrapes ( Fig. 6 View Fig C–F; Ng, 1991: fig. 6D–G). The G1 terminal segment of A. terrapes is also distinct from all congeners in that it is gently curved upwards ( Ng, 1991: fig. 6D–G). The G1 of A. rhadamanthysi is not known because no males of this species have been collected as yet.
The live colours of A. melanippe and A. kadamaiana are not known. Arachnothelphusa terrapes has brown to reddishbrown carapace, chelipeds, and walking legs ( Ng, 1991: figs. 4, 5 [erroneously not labelled in original paper]) while A. rhadamanthysi has a straw yellow coloured carapace and pereopods ( Ng & Goh, 1987: 326). The live colour of the carapace and pereopods of A. merarapensis is bright purple ( Fig. 3A, B View Fig ).
The taxonomy of genus Arachnothelphusa has been discussed at length by Ng (1991). One poorly known Sarawakian species provisionally included in the genus by Ng (1991), Parathelphusa (Liothelphusa) nobilii Colosi, 1920 , was later shown to be a species of Stygothelphusa Ng, 1989 (see Ng & Álvarez, 2000; Ng, 2013). The new species, A. merarapensis , described here is clearly member of Arachnothelphusa because it has long ambulatory legs, a transversely ovate carapace, convex anterolateral margins, rugose antero- and posterolateral carapace regions, a slender G1 whose terminal segment is elongated and at least a quarter the length of the subterminal segment, and an elongate G2 with a short distal segment ( Ng, 1991: 2).
All species of Arachnothelphusa are semi-terrestrial in habit and have been recorded from limestone caves and high elevation habitats ( Ng, 1991). Although a specimen of one species, A. terrapes , was found on a tree stump ( Ng, 1991: 11), until the present report there has been no evidence that these freshwater crabs are arboreal and live in tree holes. Arachnothelphusa merarapensis , new species, is therefore the first species of this genus known to be a true phytotelm crab. All known Arachnothelphusa species seemed to display a discrete distribution, but future molecular studies are needed to enhance our knowledge on their phylogeographic pattern in this region.
Habitat. The habitat of A. merarapensis is a steep primary dipterocarp forest with a closed canopy, near the thermal springs area of the Merarap Hot Spring Resort. The holotype male was caught from a water-filled tree-hole, ca. 22 cm deep, ca. 15 cm in diameter, and ca. 100 cm above ground ( Fig. 4 View Fig ). The bottom layer of the tree-hole consists of finely decomposed organic material (plant and arthropod debris), with the other two-thirds filled with water which increases after heavy rain. The two other specimens were caught from the water-filled crevices of tree buttresses. One buttress-crevice had a diameter of about 26 cm, was 60 cm deep, and 90 cm above the ground, and contained tightly compacted roots and leaf litter, with a hole dug into this that was apparently constructed by the crab. The other buttress-crevice runs along the side of the buttress and is about 30 cm above ground, shallow (15 cm), horizontal, and 30 cm deep. For two consecutive nights, the first two authors examined more than 15 tree-holes and crevices of different form, size, and conditions (either water-filled, dry, or debris-filled tree-holes), but we were only able to locate three specimens. This species is nocturnal and highly sensitive to light. We did not find any burrows on the soil around the tree-holes, suggesting the crab is a tree-hole specialist. Other true freshwater crabs from the Old World showing similar phytotelmic behaviour are Potamonautes raybouldi Cumberlidge & Vannini, 2004 (Potamonautidae) in Tanzania, East Africa; Globonautes macropus ( Rathbun, 1898) (Potamonautidae) in Liberia, West Africa; Malagasya goodmani ( Cumberlidge, Boyko & Harvey, 2002) (Potamonautidae) in Madagascar; and Perbrinckia scansor ( Ng, 1995) (Gecarcinucidae) in Sri Lanka ( Ng, 1995; Cumberlidge & Sachs, 1991; Bayliss, 2002; Cumberlidge et al., 2002, 2005; Cumberlidge & Vannini, 2004; Bahir et al., 2008). In Sarawak, the sesarmid crab Scandarma splendidum has been reported climbing forest trees at night in western Sarawak ( Naruse & Ng, 2007), but it is not known to be a phytotelm crab.
Conservation status. Considering unique habitat and probable limited distribution, A. merarapensis will easily be threatened by human activities, especially logging. The type locality is not protected but the forest is currently safe because it is part of the Merarap Hot Spring Resort. However, the forest in the vicinity is part of a concession allocated to logging companies and the future of the species is therefore uncertain. As such, we recommend that A. merarapensis should at least be listed as “Vulnerable” in the IUCN Red List of Threatened Species (cf. Cumberlidge et al., 2009).




| ZRC |
Zoological Reference Collection, National University of Singapore |
| SBC |
University of California |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |