
Boreocanthon Halffter, 1958
|
publication ID |
https://doi.org/ 10.5281/zenodo.7399212 |
|
publication LSID |
lsid:zoobank.org:pub:74CC7BA2-4E8D-4780-BB1B-E47370CBB19D |
|
DOI |
https://doi.org/10.5281/zenodo.7469715 |
|
persistent identifier |
https://treatment.plazi.org/id/B84F8792-9313-7F75-B1FC-B79DFA94B787 |
|
treatment provided by |
Felipe |
|
scientific name |
Boreocanthon Halffter, 1958 |
| status |
|
Genus Boreocanthon Halffter, 1958 View in CoL , restored generic status
Boreocanthon Halffter 1958: 208 View in CoL (as genus)
Canthon View in CoL ( Boreocanthon Halffter View in CoL ) (downgraded to subgenus of Canthon Hoffmannsegg View in CoL per Halffter and Martínez 1977: 81)
Type species. Ateuchus ebenus Say, 1823 View in CoL , by original designation (as Canthon ebenus [Say]).
Summary diagnosis. North American dung beetles, ball-rolling species attracted to a wide range of animal feces; usually dark or black in color, small-sized (length usually 4–7 mm); head margin conspicuously quadri- or sexdentate; parameres compressed laterally, truncate apically, lacking elongate projection of lower apical angle; hind femora lacking fine carina along anterior edge; metatibia with single apical spur.
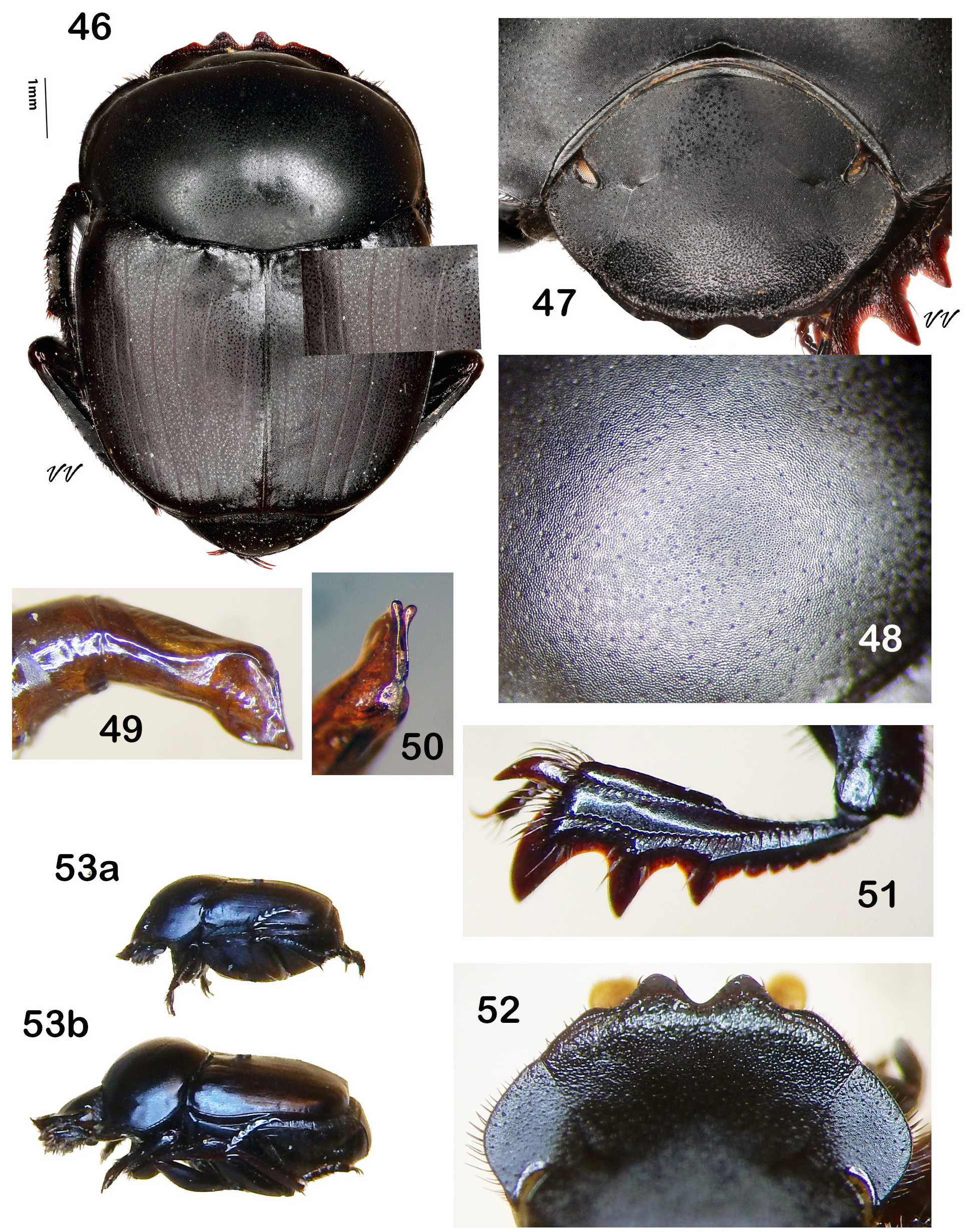
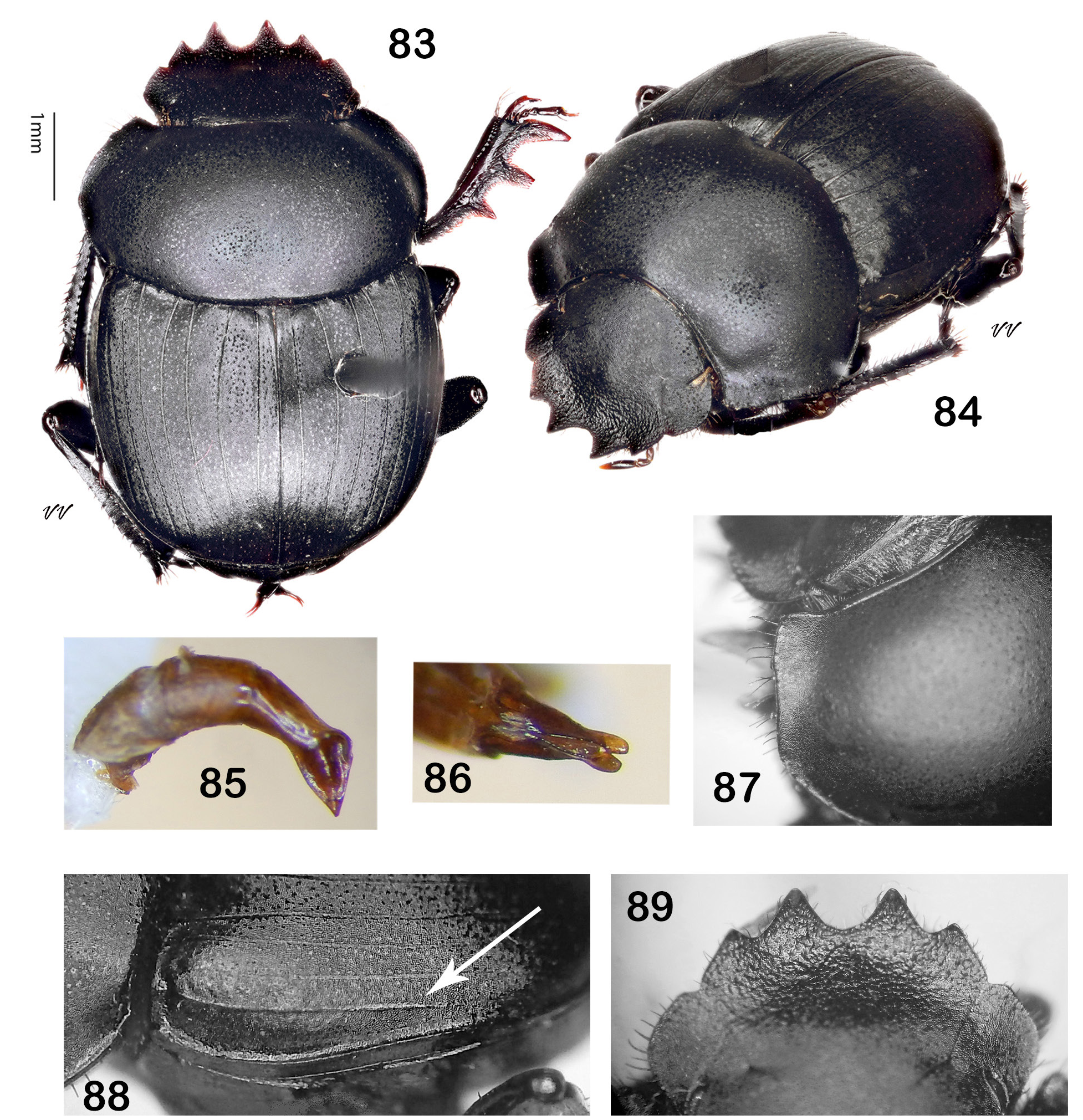
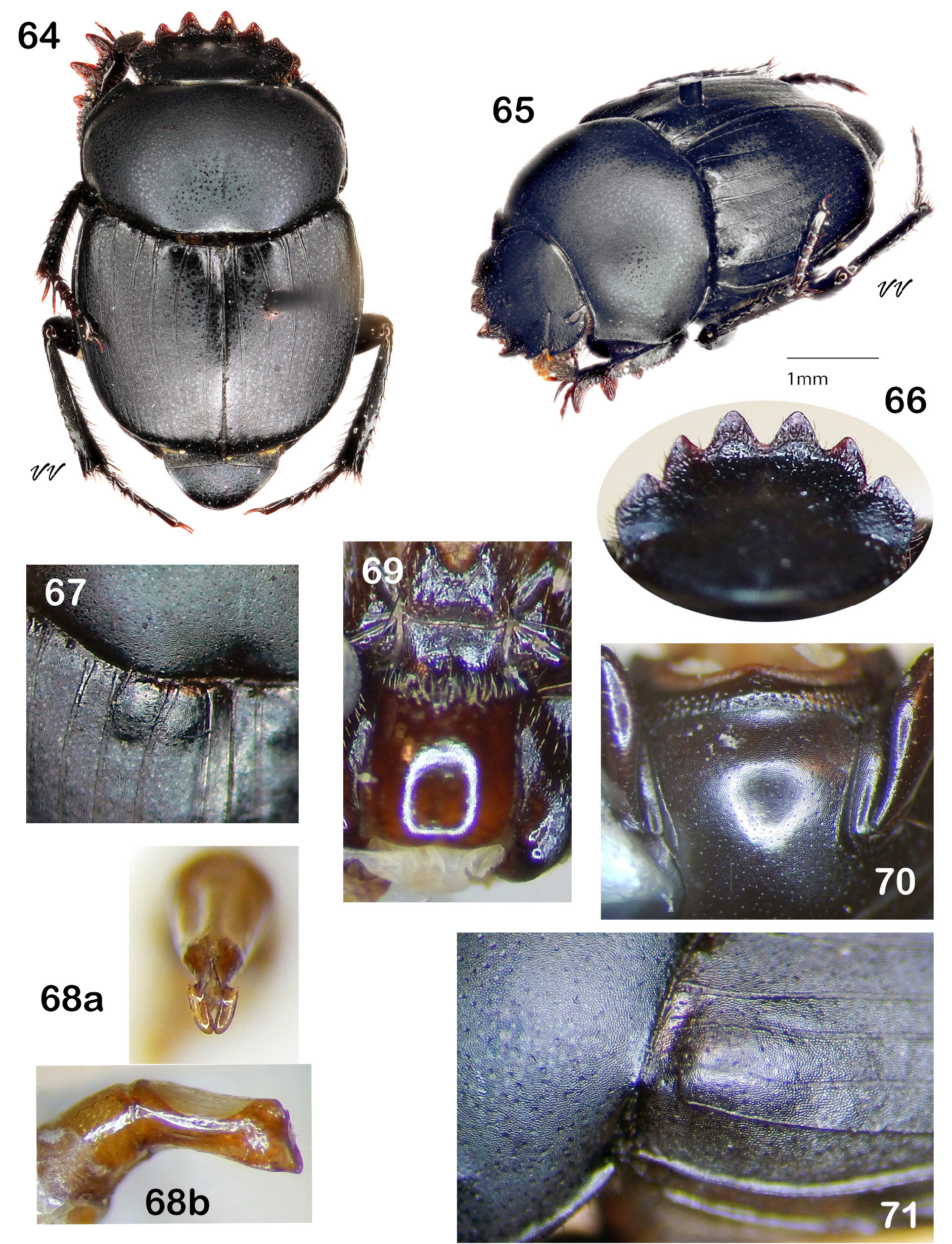
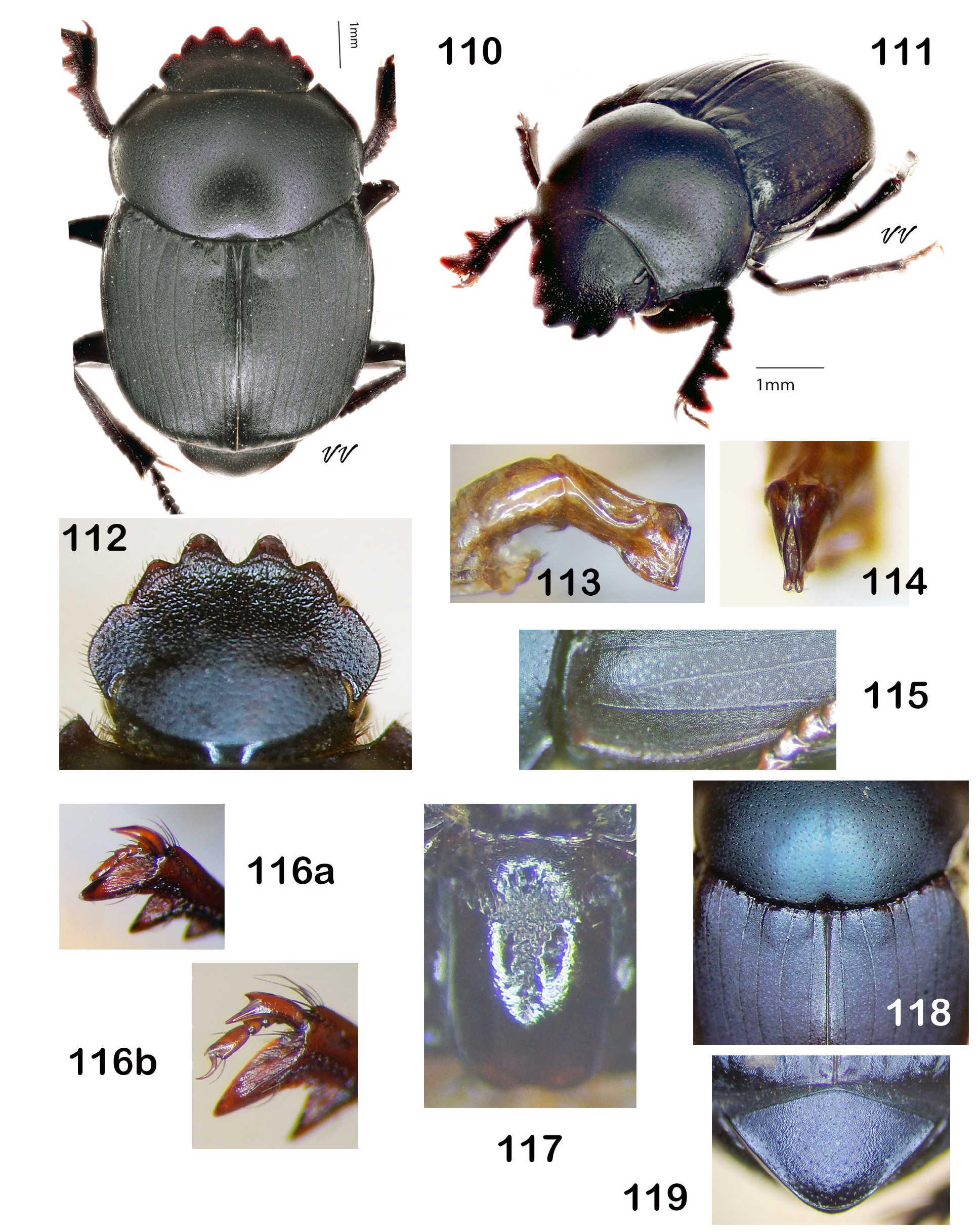
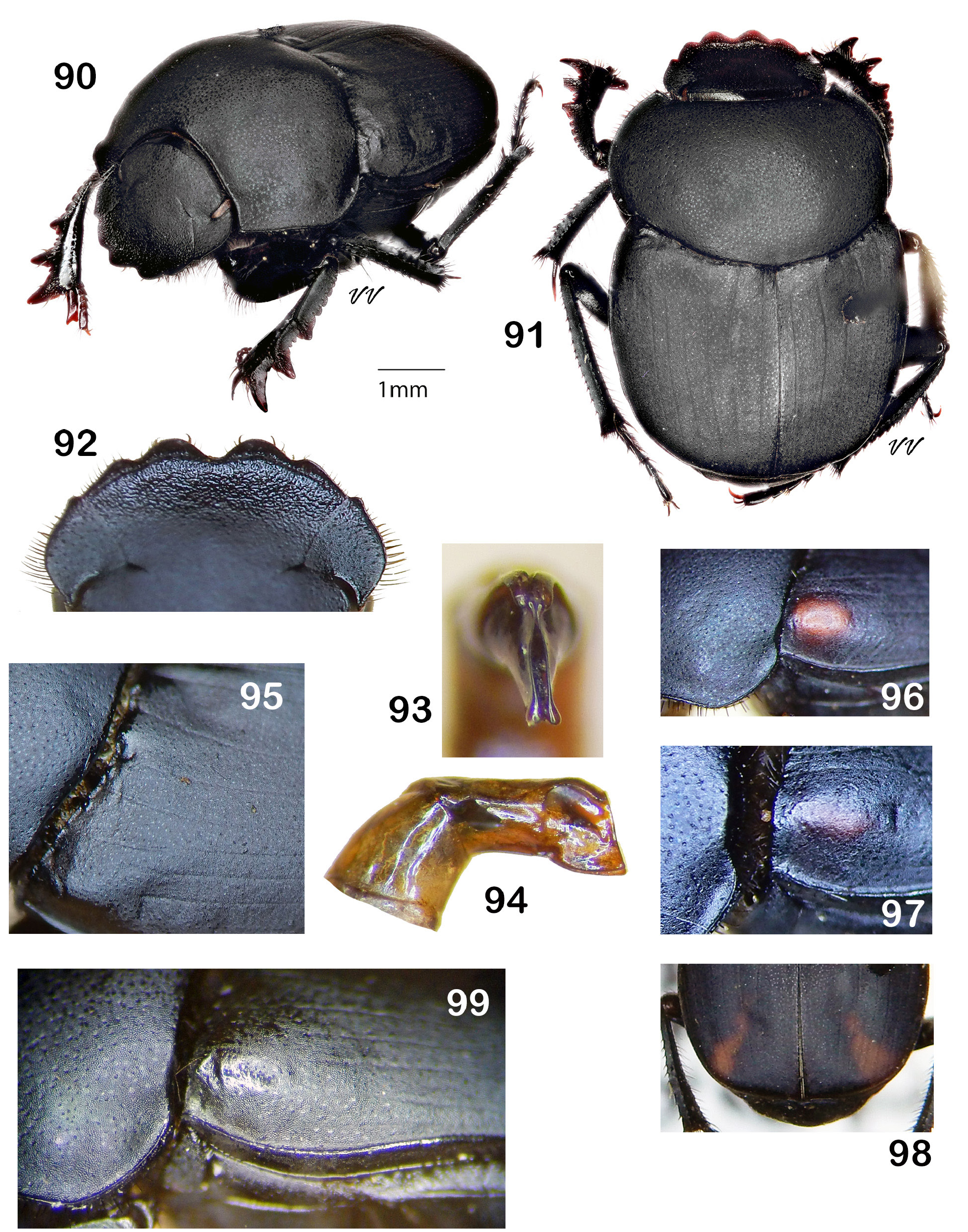
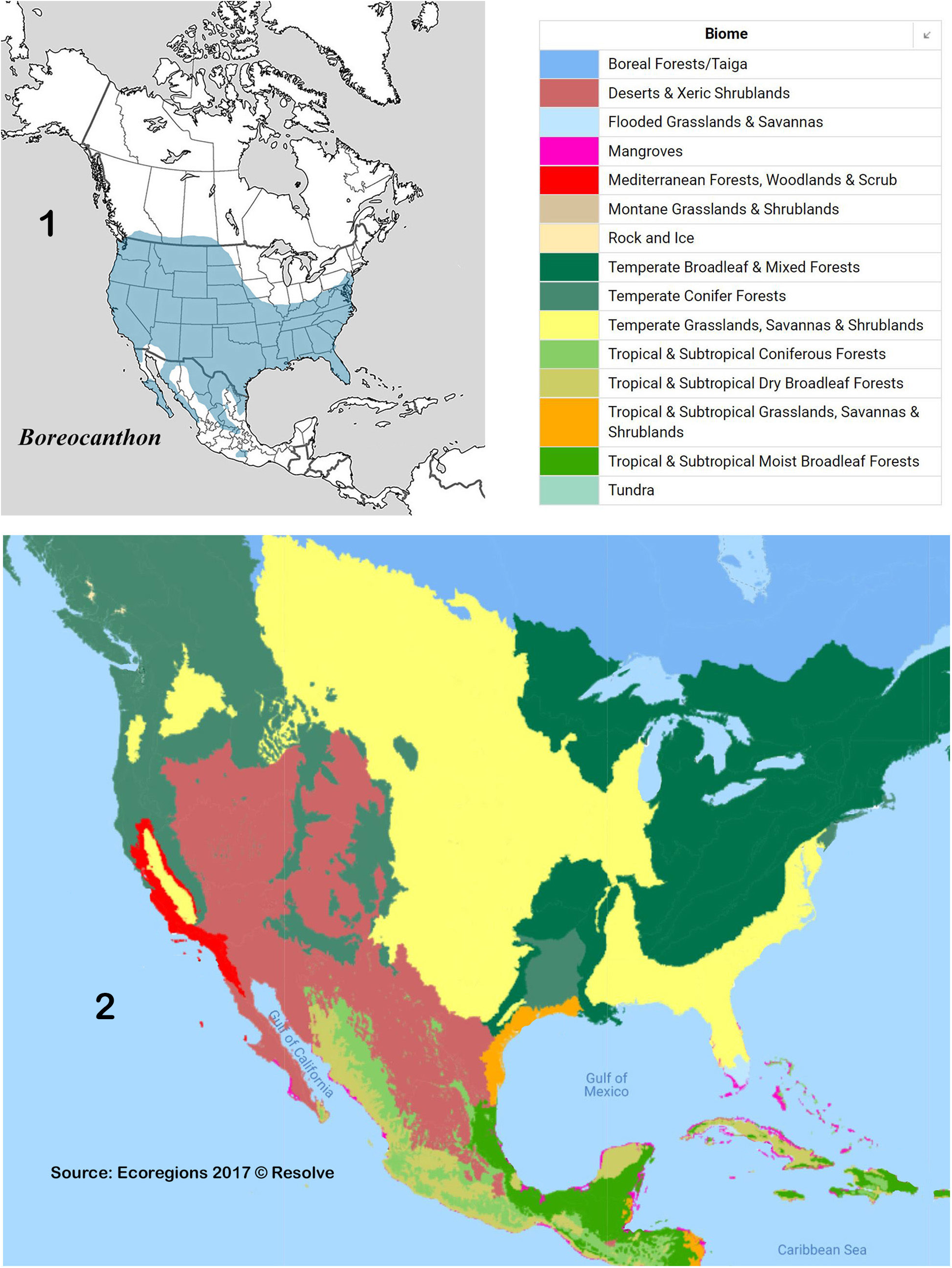
Diagnosis. Dorsum (head + pronotum + elytra + pygidium) shagreened (alutaceous), with either punctures ( Fig. 38 View Figures 36–44 , 48 View Figures 46–53 , 75 View Figures 72–81 , 105 View Figures 102–109 ) or raised granules ( Fig. 8. 19 View Figures 3–9 View Figure 10 View Figures 11–18 View Figures 19–27 ), punctures often accompanied by field of faintly lustrous microspotting ( Fig. 61 View Figures 54–62 , 88 View Figures 83–89 ); pterothoracic and abdominal sclerites shagreened, usually with accompanying flattened granules or punctures. Head: Clypeus quadridentate ( Fig. 20 View Figures 19–27 , 66 View Figures 64–71 , 79 View Figures 72–81 , 112 View Figures 110–119 ) (sometimes virtually bidentate in B. coahuilensis , Fig. 47 View Figures 46–53 ); paraocular areas often also angulate such that head appears sexdentate ( Fig. 66 View Figures 64–71 , 79 View Figures 72–81 ). Clypeal teeth reflexed, surface behind concave, finely rugose and often somewhat shinier than rest of head surface, which is usually punctate on shagreened background. Clypeal process a low, transverse ridge. Labiogular fimbria always setose, often V-shaped, usually with associated field of coarse punctures at midline ( Fig. 60 View Figures 54–62 , 69 View Figures 64–71 ). Prothorax: Subapical tubercle beneath anterior angle of pronotum usually absent. Hypomeral carina usually absent ( Fig. 24 View Figures 19–27 ), sometimes partially developed but never long, extending at most only one-fifth to one-quarter of distance to edge of prothorax ( Fig. 40 View Figures 36–44 , 108 View Figures 102–109 , arrow). Legs: Protibia strongly tridentate, usually serrate along lateral margin and between large teeth, inner margin smoothly curved ( Fig. 104 View Figures 102–109 ) or offset at level of basal tooth ( Fig. 51 View Figures 46–53 ). Anterior tibial spurs sexually dimorphic in most species: acute apically in female ( Fig. 116a View Figures 110–119 ), unequally bifurcate apically in male ( Fig. 116b View Figures 110–119 ); apex acute in both sexes in B. ebenus and B. depressipennis ( Fig. 16 View Figures 11–18 ). Posterior femora not margined anteriorly. Posterior tibiae with single apical spur. Elytra: Nine elytral striae; striae superficial, not impressed, lateral margins sometimes sharpened by fine carinulae ( Fig. 61 View Figures 54–62 , 71 View Figures 64–71 ); 9 th (epipleural) stria, if distinct, effaced anteriorly; 8 th (subhumeral) sometimes carinulate ( Fig. 88 View Figures 83–89 , arrow); 7 th and 8 th sometimes almost effaced anteriorly ( Fig. 99 View Figures 90–99 ). Interstriae always shagreened, either microspotted or granulate, sometimes presenting distinct humeral puncturing; anterior ends of first elytral interstriae depressed below level of second interstriae; anterior ends of 2 nd and 3 rd interstriae sometimes swollen ( Fig. 67 View Figures 64–71 , 111 View Figures 110–119 ), otherwise discal interstriae flattened. Genital capsule: Distal portion of parameres laterally compressed, truncate, posterior edges straight, appressed, profile varies from abbreviated ( Fig. 113 View Figures 110–119 ) to elongate triangle ( Fig. 85 View Figures 83–89 ), lacking any curved elongation of the lower apical angle (as in Fig. 43a View Figures 36–44 ). General: Small, length rarely over 7.5 mm, usually 4.5–6 mm. North America from Trans-Mexican Volcanic Belt in southern Mexico, through the United States into far south-central Canada ( Fig. 1 View Figures 1–2. 1 ).
Comments. Boreocanthon is here restored to its original status of genus because, like its close relative, Melanocanthon , it exhibits a distinguishing suite of morphological features (see Diagnosis) and a plainly North American distribution. I have not discovered any unique feature (synapomorphy) that both unites its 13 species and separates them from other “ Canthon ,” sensu lato, and, therefore, I cannot argue with a high degree of confidence that the group is monophyletic. Boreocanthon and Melanocanthon , along with the pilularius species-group of Canthon (sensu Halffter 1961) constitute the endemic United States contingent of the North American fauna of deltochiline Scarabaeinae . All other deltochilines occurring in temperate North America, north of the Rio Grande, are recently arrived representatives of southern groups; these include Deltochilum gibbosum (Fabricius) , Pseudocanthon perplexus (LeConte) , Canthon (Glyphrocanthon) viridis (Palisot de Beauvois) , C. (Canthon) humectus (Say) , C. (C.) indigaceus LeConte and C. (C.) cyanellus LeConte.
Boreocanthon is closely related to Melanocanthon , and the two are sometimes confused in collections. The latter group, however, is distinguished by having two apical spurs on the hind tibia (rather than one) as well as, among other features, subtle differences in dorsal sculpturing and the shape of the head and sides of the prothorax and elytra, and a distribution largely confined to the southeastern quadrant of the United States, including the eastern one-half of Texas. There are presently four recognized species of Melanocanthon , but my informal impression is that there may be but three. A future, more rigorous look at Melanocanthon is very much in order. Halffter et al. (2022) recently created the subgenus Bajacanthon to accommodate the unique Baja California endemic species, Canthon obliquus Horn. In their phylogenetic analysis ( Melanocanthon was not included) the new genus emerged as sister group to Boreocanthon , with which it shares several features, especially the form of the parameres ( Fig. 44a,b View Figures 36–44 ).
Boreocanthon View in CoL is a Nearctic taxon with deep Neotropical roots. The center of diversity of the genus is clearly the United States and adjacent northern Mexico, where it undoubtedly originated. It, as well as Melanocanthon View in CoL and the pilularius View in CoL species group of Canthon View in CoL , are the only representatives of the Neotropical tribe Deltochilini View in CoL that originated and diversified in the Nearctic. The historical biogeographic perspective proposed by Kohlmann and Halffter (1990) seems reasonable in the context of the results of this study: “The ancestor of Melanocanthon View in CoL - Boreocanthon View in CoL would have been distributed in northern Mexico and south-central USA, while the area was covered by tropical deciduous forest. By the end of Miocene the forest started to be replaced by grasslands and piñon-oak woodland, a process that was advanced in the Pliocene by the spread of pine forest and very recently the appearance of deserts. Most Boreocanthon species … live in this area, which leads us to think that this was the main evolutionary arena for this group. Nowadays, many of the species live in grasslands of the Great Plains … or in arid zones …” (p. 13). In the context of Gonzalo Halffter’s theory of the Mexican Transition Zone ( Morrone 2015; Halffter 2017; Halffter and Morrone 2017 and numerous references therein), the modern Nearctic taxa, including Boreocanthon View in CoL , are the descendants of deltochilines among a wave of Neotropical immigrants that arrived in North America during early Cenozoic, where they originated and began to radiate during Miocene-Pliocene times. This early wave of taxa from the south produced the founding cenocron of Halffter’s Mexican Plateau Dispersal Pattern, one of five distribution patterns comprising the biogeographical panorama of the Mexican Transition Zone ( Halffter and Morrone 2017). This cenocron included many other scarabaeines and other insects, including Phanaeus View in CoL , whose biogeographical history in part parallels that of the Boreocanthon View in CoL and other Nearctic deltochilines ( Kohlmann and Halffter 1990; Halffter and Morrone 2017) as well as other groups ( Edmonds 1994).
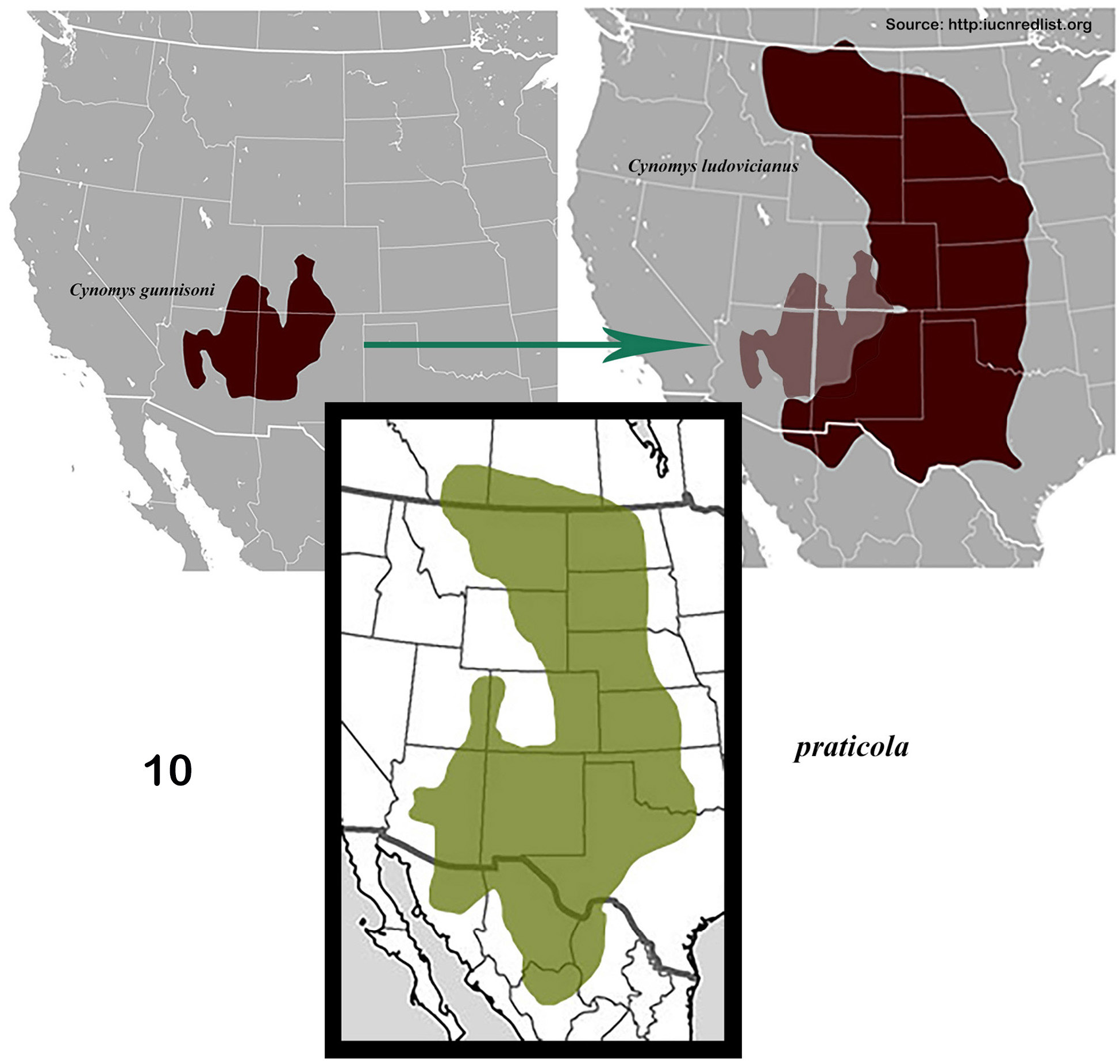
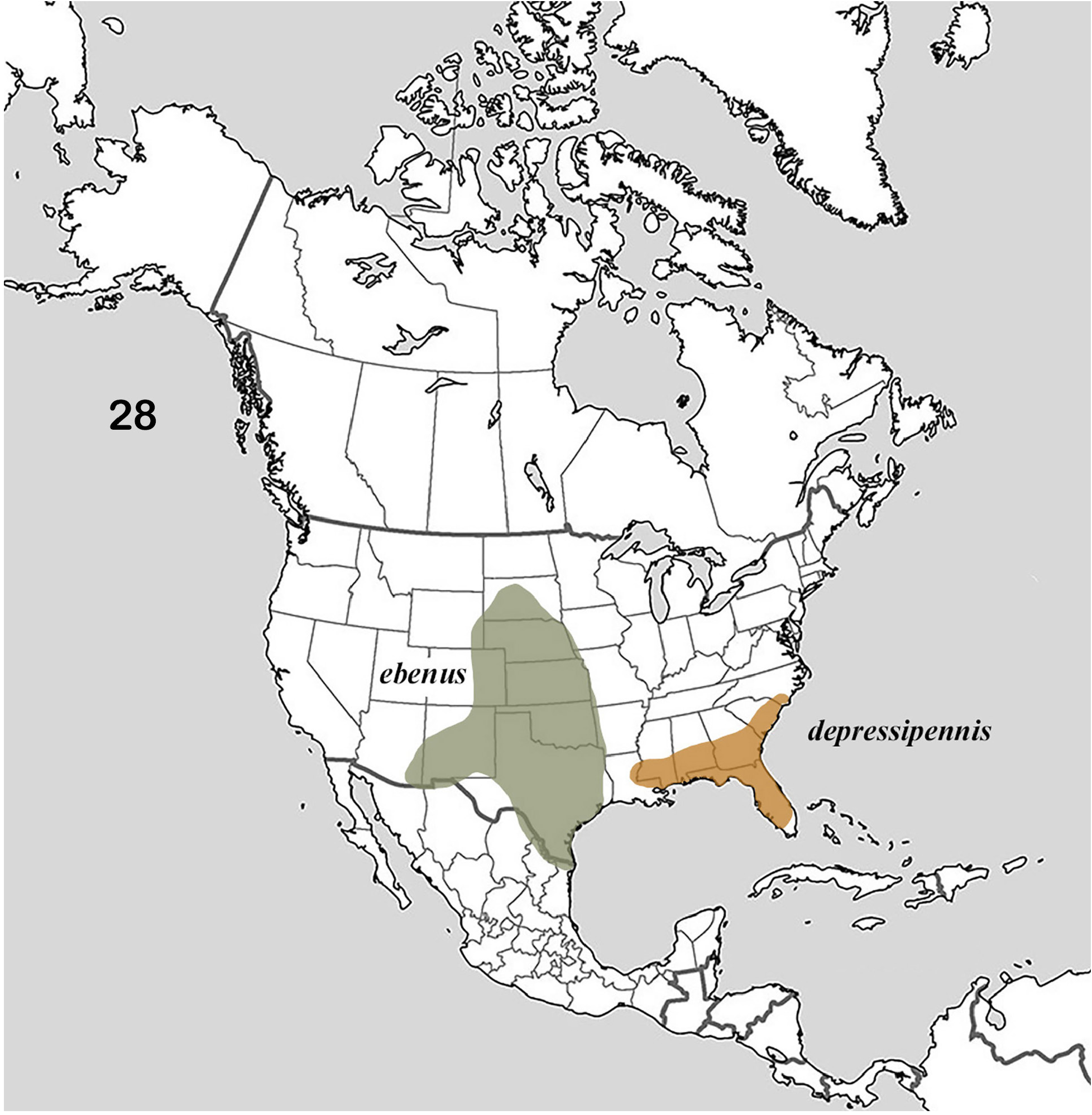
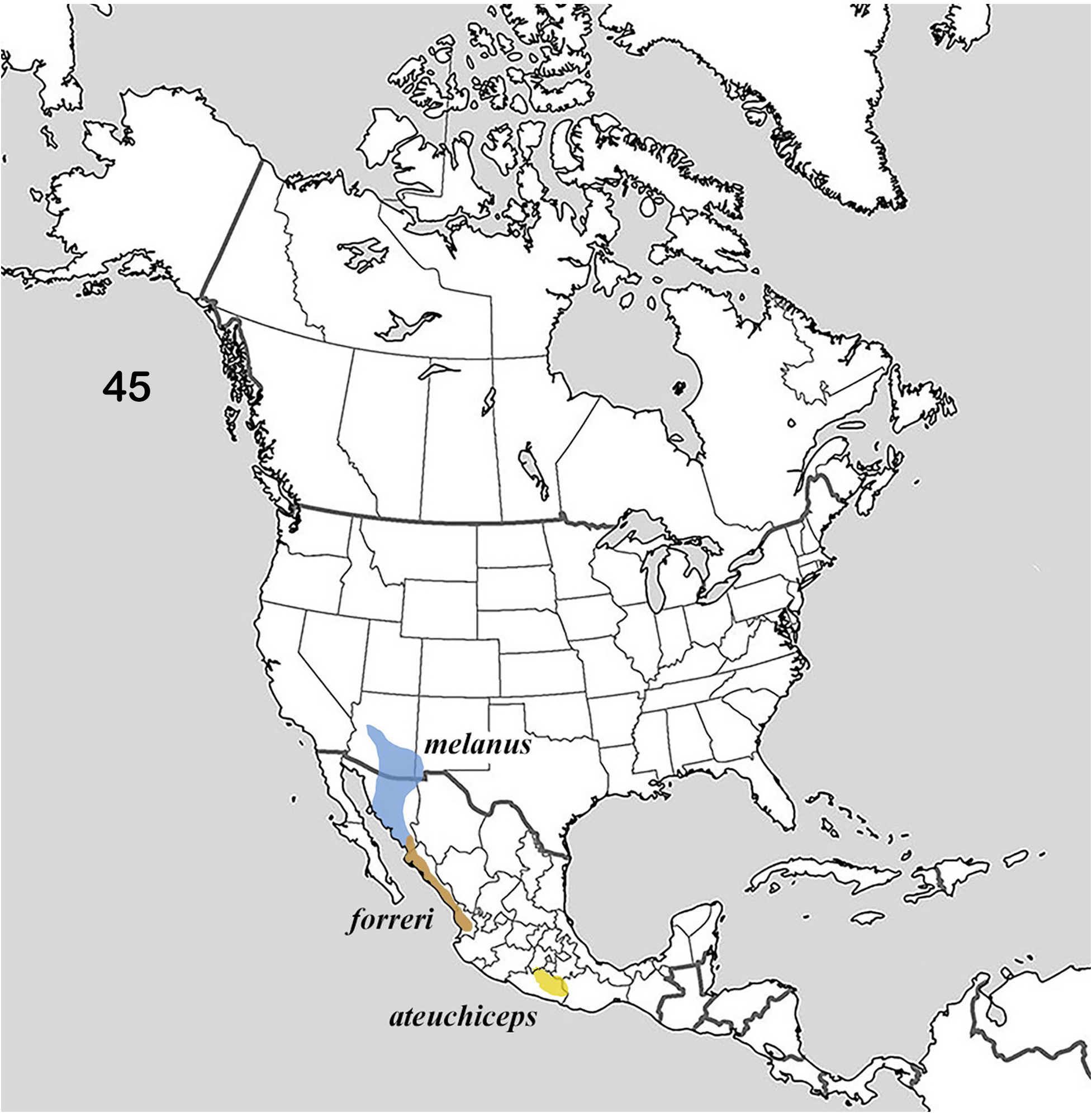
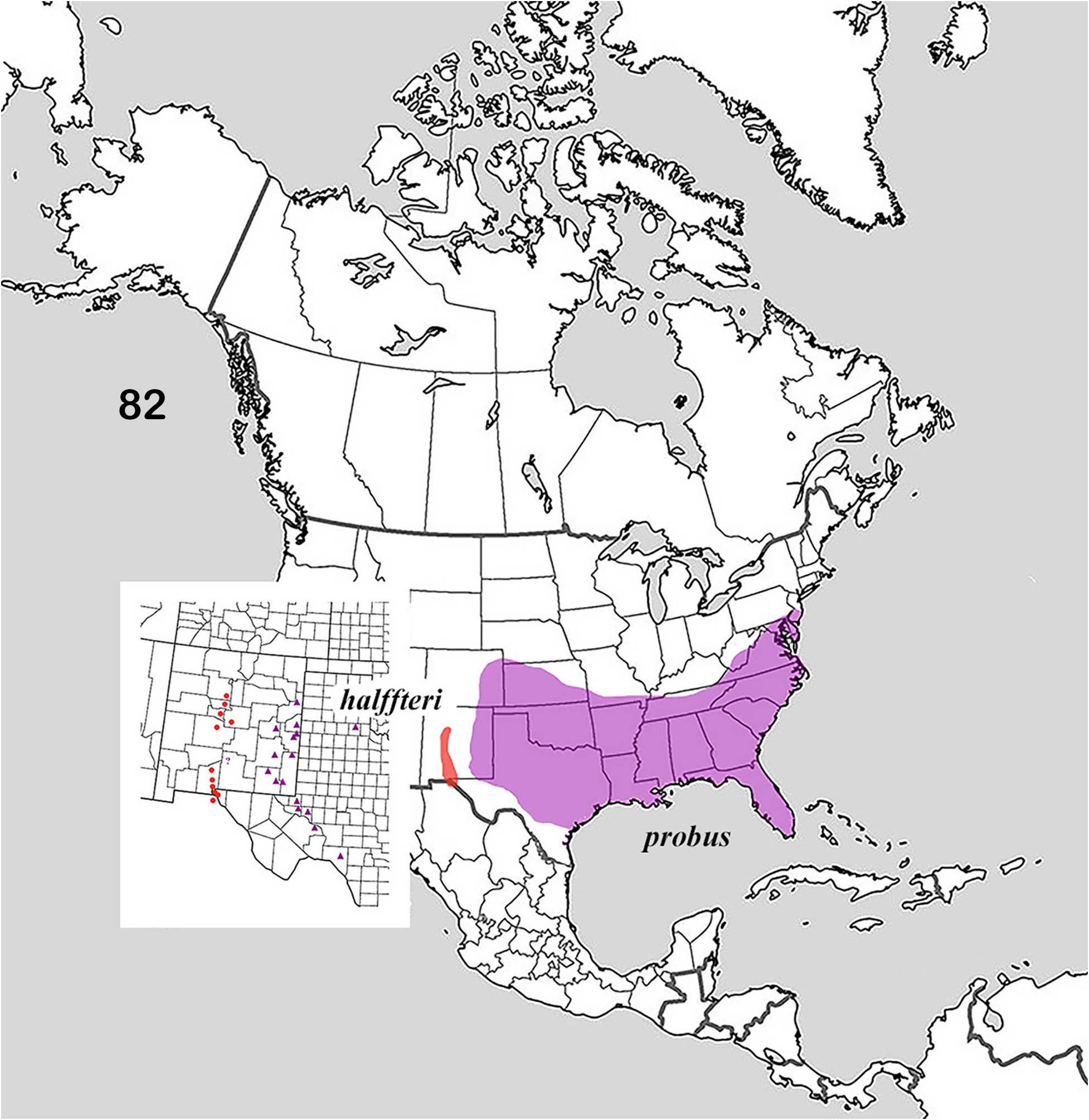
The distribution of the genus covers much of North America ( Fig. 1 View Figures 1–2. 1 ). Of interest is the fact that the distributions of the member species of putative species pairs (B. ebenus-B. depressipennis , Fig. 28 View Figure 28 ; B. melanus-B. forreri , Fig. 45 View Figure 45 ; B. puncticollis-B. integricollis , Fig. 63 View Figure 63 ; B. probus-B. halffteri , Fig. 82 View Figure 82 ; and B. simplex-B. coahuilensis , Fig. 100 View Figures 100–101 ) are allopatric or nearly so, which suggests a long history of vicariant/dispersal events punctuating the history of the group (see, for example, Gámez et al. 2016). The ecogeographic distributions of Boreocanthon species conform rather closely to the vegetation biomes depicted in Figure 2 View Figures 1–2. 1 : B. depressipennis and most B. probus , to eastern coniferous open-canopy forests; B. praticola and B. ebenus , to mid-latitude steppe and prairie; B. lecontei and B. integricollis to arid south Texas shrubland; and B. simplex to the complex highly variable mosaic of biomes present west of the Rocky Mountains. Western populations of B. probus , as well as B. halffteri , have colonized west Texas / New Mexico steppe and desert shrub habitats. All the Mexican species ( B. melanus , B. forreri , B. ateuchiceps , B. coahuilensis , and B. puncticollis ) are associated with desert and desert shrub areas.
That the diversity hotspot for Boreocanthon lies in the southwestern portion of the United States is unsurprising in that it is the focus of diversity for numerous vertebrate and invertebrate groups (see Parmenter et al. 1995). In this complex assemblage of arid and semi-arid range and basin habitats characteristic of western Texas, and southern New Mexico and Arizona, seven species come together: B. puncticollis , B. melanus , B. ebenus , B. halffteri , B. simplex , B. praticola and B. probus . Only two, however, are endemics: B. halffteri and B. melanus . For comparison, in the United States west of the Rocky Mountains north of about 34° latitude, an area much larger than the U.S. Southwest, is home to a single, albeit highly variable species, B. simplex . All other species, B. lecontei , B. integricollis , B. depressipennis , B. coahuilensis , B. ateuchiceps and B. forreri can be considered peripheral, relatively localized relatives of these seven. What this says about the biogeographical history of the genus is ripe for analysis.
Available data bespeak a group that is fundamentally coprophagous, with but very few records of attraction to carrion and none to fungi or other decomposing organic matter. Available information suggests the group to be almost exclusively coprophagous but highly generalist in food choice. Reported dung sources include a wide variety of wild and domestic animals: humans, hog, cattle, bison, horses, burros, deer, elk, rabbit, prairie dog, fox, bear, coyote, dog, domestic cat, javelina (collared peccary), opossum, chicken. In only one known instance does a species exhibit a preference for a particular food source: B. praticola (q.v.) for dung pellets of prairie dogs ( Cynomys spp. ). Zunino and Halffter (2007) report several casual associations between species of Boreocanthon and various vertebrate nests: B. simplex with ground squirrel ( Citellus ) burrows; B. puncticollis with packrat ( Neotoma ) and kangaroo rat ( Dipodomys ) nests. Presumably, all Boreocanthon conserve the ball-making/rolling (telocoprid) strategy characteristic of deltochilines (see Tonelli 2021). While there are some anecdotal observations that they do, I know of no formal corroborative studies on the feeding and nesting behaviors of any species of Boreocanthon .
Pollock and Lavigne (2019) reported that B. praticola , along with Digitonthophagus gazella (Fabricius 1787) and Canthon floridanus Brown, 1946 (until recently known as C. imitator Brown ; see Génier 2019) are regular prey items for various species of robber flies ( Diptera : Asilidae ) in eastern New Mexico.
Robinson (1948) presented the first modern key to the species of this genus. He included the ten thenknown species of Boreocanthon in the United States and his study heretofore has been the standard taxonomic reference for the group. Boreocanthon species group in his key to Canthon (couplets [0] through 2) on the basis of the following combination of characters: a) no [complete] hypomeral carina; b) anterior edge of hind femora not bearing a carina; and c) hind tibia with a single apical spur. As defined here, Boreocanthon embraces the thirteen valid species listed below. A total of 22 available and one unavailable species-group names, and their status as valid or otherwise, are assigned to the genus and listed below (original generic placement in parentheses; brackets enclose reference to designator; valid names and new nomenclatural actions are in bold type):
abrasus LeConte 1859c: 11 (Canthon) View in CoL —jr. subj. syn. probus Germar View in CoL [ Horn 1870: 45]
antiguus Pierce 1946: 120 ( Canthon View in CoL )—jr. subj. syn. simplex LeConte View in CoL [ Miller et al. 1981:626]
ateuchiceps Bates 1887: 35 (Canthon) View in CoL —valid name
bisignatus Balthasar 1939: 229 (Canthon) View in CoL —jr. subj. syn. militaris Horn, new synonymy
coahuilensis Howden 1966: 732 (Canthon) View in CoL —valid name
corvinus Harold 1868: 129 (Canthon) View in CoL —jr. subj. syn. simplex LeConte View in CoL [ Horn 1870: 46]
depressipennis LeConte 1859c: 11 (Canthon) View in CoL —valid name
ebenus Say 1823: 208 (Ateuchus) —valid name ebeneus Say 1823: 208 (Ateuchus) View in CoL —suppressed (see Comments for ebenus )
forreri Bates 1887: 31 (Canthon) View in CoL —valid name
halffteri Edmonds (Boreocanthon) View in CoL — new species
humeralis Horn 1870: 46 ( Canthon View in CoL )—jr. subj. syn. simplex LeConte View in CoL [ Robinson 1948: 87]
integricollis Schaeffer 1915: 50 (Canthon) View in CoL —valid name
lecontei Harold 1868: 68 (Canthon) View in CoL —valid name
melanus Robinson 1948: 88 (Canthon) View in CoL —valid name
militaris Horn 1870: 46 ( Canthon View in CoL )—jr. subj. syn. simplex LeConte View in CoL [ Robinson 1948: 87]
mixtus Robinson 1948: 91 (Canthon) View in CoL —jr. subj. syn. puncticollis LeConte View in CoL , new synonymy
nyctelius Bates 1887: 31 (Canthon) View in CoL —jr. subj. syn. puncticollis LeConte View in CoL [ Schaeffer 1915: 50]
praticola LeConte 1859c: 10 (Canthon) View in CoL —valid name
probus Germar 1823: 98 (Ateuchus) View in CoL —valid name
puncticollis LeConte 1866: 381 (Canthon) View in CoL —valid name
simplex LeConte 1857: 41 (Canthon) View in CoL —valid name
vetustus Pierce 1946: 122 ( Canthon View in CoL )—jr. subj. syn. praticola LeConte View in CoL [Robinson 1981: 626]
Key to the Species of Boreocanthon Halffter View in CoL
Note. Boreocanthon View in CoL comprises species that are often widely distributed and patently variable. The following key relies mostly upon generalizations about structure and, therefore, cannot be expected to accommodate inevitable exceptions. Reliability of identifications will be enhanced if target specimens are free or largely so of significant wear, especially to the head margin and to the dorsal surface. Since closely related Boreocanthon species are commonly allopatric, accurate locality information is an important corroborator of diagnostic structural features in this genus. The text page position for each species treatment is indicated in brackets following the species name.
1. Dorsal sculpturing (including the pygidium) consisting of coarse granulation on shagreen (alutaceous) background ( Fig. 8 View Figures 3–9 , 13 View Figures 11–18 , 27 View Figures 19–27 ); granules discrete, round, raised above surface, distinct punctures and shiny microspots lacking. Notch separating clypeus from paraocular area large, setting off tooth-like anterior angle of paraocular area such that head appears strongly sexdentate ( Fig. 4 View Figures 3–9 , 11 View Figures 11–18 , 20 View Figures 19–27 )............... 2
— Dorsal sculpture variable but not including discrete, raised, rounded granules; pronotum either punctate on shagreen background, or ( B. lecontei View in CoL , Fig. 31 View Figures 29–35 ) covered by dense, fine granulation largely supplanting shagreen background. Dorsum ( Fig. 48 View Figures 46–53 , 75 View Figures 72–81 ) usually presenting shiny microspots, especially on pronotum (except B. lecontei View in CoL ), elytra and often also on head and pygidium. Paraocular areas and dentition variable................................................................................ 4
2. Form of protibial spur same in both sexes, viz, strongly curved laterad and tapering to acute point ( Fig. 20 View Figures 19–27 ). Posterior portion of lateral pronotal margin asperate (serrate). Hypomeral carina absent ( Fig. 24 View Figures 19–27 ). Protibia (viewed along inner margin) abruptly widened (offset) at level of third (basal) tooth; inner apical angle produced as small, conical, apically-directed tooth ( Fig. 16 View Figures 11–18 , arrow). Pygidium distinctly impressed basally, resulting transverse concavity often divided by median, longitudinal swelling ( Fig. 14 View Figures 11–18 , 27 View Figures 19–27 ). Compressed tip of parameres (viewed laterally) in form of elongate triangle ( Fig. 18 View Figures 11–18 ).................. 3
— Protibial spur sexually dimorphic: strongly curved laterad and tapering to acute point in female; nearly straight to curved laterad, apically slightly expanded and obliquely truncated or asymmetrically notched in male (as in Fig. 116a,b View Figures 110–119 ). Posterior portion of lateral pronotal margin smooth. Hypomeral carina present, sometimes only slightly developed. Protibia usually only gently sinuous at level of basal tooth. Pygidium evenly convex, not impressed basally ( Fig. 6 View Figures 3–9 ). Compressed tip of parameres short, truncated.
Length 5–10.5 mm. Very widely ranging from south central Canada to northern Mexican Plateau, extending westward into eastern Arizona ( Fig. 10 View Figure 10 )....... Boreocanthon praticola (LeConte) View in CoL [p. 13]
3. Pronotal granules uniform in size and shape, evenly and densely distributed and seldom coalescent ( Fig. 11, 13b View Figures 11–18 ). Surface granulation extending to pteropleura but much weaker on median area of metaventrite and absent or only weakly evident on abdominal ventrites. Circumnotal ridge becoming very fine posteriorly, extending around posterior pronotal angle mesally to point adjacent to 7 th or 8 th elytral stria ( Fig. 13b View Figures 11–18 , arrows). Dorsum dark but almost always presenting metallic green coloration (viewed under bright light). Length 6–10 mm. Southeastern United States from Louisiana to North Carolina ( Fig. 28 View Figure 28 )..................................................... Boreocanthon depressipennis (LeConte) View in CoL [p. 18]
— Pronotal granules larger, variable sized and frequently coalescent, especially on disk ( Fig. 13a View Figures 11–18 , 19 View Figures 19–27 ). Surface granulation extending to pteropleura, at least anterior portion of median area of metaventrite, and at least sides of all abdominal ventrites. Circumnotal ridge ending at posterior pronotal angle, not extending onto posterior pronotal margin ( Fig. 13a View Figures 11–18 ). Black, only very rarely with any hint of color. Length 6–11 mm. United States west of Mississippi River to Rocky Mountains with southwestern extension through Texas to southeast Arizona ( Fig. 28 View Figure 28 )........................... Boreocanthon ebenus (Say) View in CoL [p. 20]
4. Pronotum very finely, densely granulate, granulation highly coalescent largely supplanting shagreen background; punctures absent ( Fig. 29–31 View Figures 29–35 ). Paraocular notch very small, sometimes almost obsolete, scarcely if at all breaching raised lateral margin of head ( Fig. 29, 32 View Figures 29–35 ). Smaller beetles, overall length not exceeding 5 mm. Southern Texas and northern Tamaulipas ( Fig. 100 View Figures 100–101 )... Boreocanthon lecontei (Harold) View in CoL [p. 26]
— Pronotum distinctly punctate, usually on distinctly shagreen background, punctures can be bold or very fine and widely dispersed such that pronotal surface appears almost smooth; pronotum never granulate. Paraocular notch variable................................................................. 5
5. Dorsum bright metallic green, often with yellow or coppery highlights ( Fig. 36–39 View Figures 36–44 ). Pronotum strongly but not densely punctate, shiny, shagreen background mostly effaced, sometimes detectable in isolated patches near anterior angles ( Fig. 38 View Figures 36–44 ). Upper surface of head strongly roughened by large, closely packed, coalesced punctures. Length 4.5–6.0 mm. Sierra Madre del Sur, Mexico ( Fig. 45 View Figure 45 )........................................................................ Boreocanthon ateuchiceps (Bates) View in CoL [p. 28]
— Dorsum dark, usually black, sometimes with blue or green highlights (producing dark, metallic green color in B. forreri View in CoL ). Pronotum punctate, often finely and sparsely so; shagreen background and shiny spots distinct, only rarely obscured. Upper surface of head smooth, with subtle punctures except anterior portion of clypeus, which is usually distinctly roughened. Length variable. Distribution north of Transverse Volcanic Range in Mexico northward to southern Canada..................................... 6
6. Paraocular notch obsolete, anterior angle of paraocular area never accentuated ( Fig. 47, 52 View Figures 46–53 ). Clypeal margin usually bearing four perceptible teeth, median pair short, broadly rounded, wider at base than long, lateral two always small and inconspicuous and sometimes virtually absent such that clypeus appears bidentate ( Fig. 46–47 View Figures 46–53 ). Pronotum (viewed in profile) strongly raised above level of elytra (“humpbacked,” Fig. 53b View Figures 46–53 ). Protibia abruptly widened, inner margin offset at level of basal lateral tooth ( Fig. 51 View Figures 46–53 ). Larger sized, length 7–11 mm. Distribution restricted to northwestern flank of Sierra Madre Oriental in southwest Nuevo Leon /southeast Coahuila, Mexico ( Fig. 101 View Figures 100–101 )................................................................................. B oreocanthon coahuilensis (Howden) [p. 31]
— Paraocular notch (in unworn specimens) evident, sometimes very small, more often excising head margin and setting off angular prominence of paraocular area; shape of head margin variable but always distinctly quadridentate or sexdentate. Pronotum (viewed in profile) most often evenly rounded and at most only slightly raised above level of elytra. Protibia only gradually widened toward apex, any offset only weakly indicated. Smaller, length generally less than 7 mm. Widely distributed in Mexico and the United States............................................................................ 7
7. Lower apical angles of parameres expanded laterally as thin rounded or elongate flanges ( Fig. 56–57 View Figures 54–62 , 68a View Figures 64–71 ). Margins of elytral striae 1–4 and sometimes also 5–6 sharply defined by carinulae extending from base at least one-third total length ( Fig. 71 View Figures 64–71 ). Paraocular notch deep, setting off strong, angulate portion of paraocular area, head appearing clearly sexdentate ( Fig. 62 View Figures 54–62 , 66 View Figures 64–71 )................................ 8
— Lower apical angles of parameres in form of elongate, distally rounded swellings ( Fig. 77 View Figures 72–81 , 86 View Figures 83–89 , 93 View Figures 90–99 ), not flanged. Margins of elytral striae either not carinulate, or only briefly so anteriorly and then in occasional specimens showing metallic highlights. Paraocular notch variable.................................. 9
8. Ventral apical angle of parameres, viewed caudally, with ear-shaped, rounded or slightly angulate flanges ( Fig. 56–57 View Figures 54–62 ). Second and third elytral interstriae conspicuously swollen anteriorly, these raised areas flanking deeply impressed first interstriae ( Fig. 59 View Figures 54–62 ). Pronotum distinctly impressed posteromedially, resulting concavity almost smooth and contiguous with depression formed by first interstriae of elytra ( Fig. 55 View Figures 54–62 ). Epipleural (9 th) stria usually visible posteriorly. Mesoventrite at most with few central punctures. Dorsum, viewed in bright light under magnification, with strong, royal blue (very rarely green) undertone (intensified when surface is wet). Length 4.0– 6.5 mm. Baja California del Sur, the Mexican Plateau and adjacent areas of Arizona, New Mexico and western Texas ( Fig. 63 View Figure 63 )...................................................................... Boreocanthon puncticollis (LeConte) View in CoL [p. 33]
— Ventral apical angle of parameres, viewed caudally, in form of elongate flanges ( Fig. 68 View Figures 64–71 ). Second and third elytral interstriae usually not conspicuously swollen; pronotum lacking distinct posteromedian concavity. Epipleural (9 th) stria effaced. Mesoventrite completely coarsely punctured ( Fig. 70 View Figures 64–71 ). Dorsum black, lacking strong blue undertone. Length 4.0– 5.5 mm. Southern Texas and far northeast Mexico ( Fig. 63 View Figure 63 )................................................. Boreocanthon integricollis (Schaeffer) View in CoL [p. 38]
9. Paraocular notch large, lateral margin of paraocular area strongly salient ( Fig. 72, 74 View Figures 72–81 ). Clypeal teeth large, acute. United States east of the Rocky Mountains and central New Mexico ...................... 10
— Paraocular notch obscure, anterior angle of paraocular area interrupting curve of head margin as small, angulate corner ( Fig. 92 View Figures 90–99 , 106 View Figures 102–109 , 112 View Figures 110–119 ). Clypeal teeth obtuse, broadly rounded apically. Southeastern Arizona southward along coast of mainland Mexico to Nayarit ................................... 11
Note. Couplet 10 separates taxa for which intermediate populations exist in west Texas and eastern New Mexico. Some specimens from Ward and Winkler Counties in Texas, and Eddy and Lea Counties View in CoL in New Mexico can be difficult to sort morphologically; they are here regarded as B. probus View in CoL on the basis of distribution. See Comments for B. probus View in CoL .
10. Pronotum not at all or only weakly (narrowly) explanate anterolaterally. Paraocular notch narrow, setting off strong, anteriorly directed tooth, head strongly sexdentate ( Fig. 72, 74 View Figures 72–81 ). Anterior portion of head behind clypeal teeth often abruptly shinier and rougher than posterior portion, which is smooth and punctate over shagreen background ( Fig. 79 View Figures 72–81 ). Subhumeral (8 th) stria simple or only weakly, if at all carinulate. Profile of compressed distal portion of parameres in form of equilateral triangular ( Fig. 76 View Figures 72–81 ). Length 4.0– 5.5 mm. Eastern United States from Atlantic coast to Rocky Mountains roughly south of 41 st parallel, Texas and New Mexico east of the Pecos River ( Fig. 82 View Figure 82 )........... Boreocanthon probus (Germar) View in CoL [p. 40]
— Pronotum widely explanate, upturned narrowly at lateral angle and progressively more widely so toward anterior angle ( Fig. 84, 87 View Figures 83–89 ). Paraocular notch setting off transverse anterior margin of paraocular area, head strongly quadridentate ( Fig. 83, 89 View Figures 83–89 ). Anterior portion of clypeus dull, only rarely weakly shinier than rest of head surface ( Fig. 89 View Figures 83–89 ). Subhumeral (8 th) elytral stria carinulate, usually sharply so from umbone to midpoint of elytral margin or slightly beyond ( Fig. 88 View Figures 83–89 , arrow). Compressed distal portion of parameres, viewed laterally, in shape of elongate triangle ( Fig. 85 View Figures 83–89 ). Length 4.5–6.5 mm. Rio Grande drainage of New Mexico and Texas from Albuquerque southward to El Paso and Chihuahua ( Fig. 82 View Figure 82 , inset)............................................. Boreocanthon halffteri View in CoL , new species [p. 45]
11. Median clypeal teeth broadly rounded, lateral clypeal teeth small, length less than half that of median teeth; margin of head appearing quadridentate ( Fig. 92 View Figures 90–99 ). Pronotal puncturing usually only weakly indicated, only occasionally conspicuous (×10). Subhumeral (8 th) elytral stria never carinulate ( Fig. 99 View Figures 90–99 ); striae 7 and 8, sometimes also 6, often noticeably weaker than discal striae, anterior one-half or more of 7 th and 8 th sometimes nearly effaced. Compressed distal portion of parameres (viewed laterally) triangular ( Fig. 94 View Figures 90–99 ). Widely distributed in the western United States and Canada from the Rocky Mountains westward to Pacific coast, extending northward to southern British Columbia and southward to northwestern Baja California ( Fig. 100–101 View Figures 100–101 ).............................. Boreocanthon simplex (LeConte) View in CoL [p. 48]
— Median clypeal teeth rounded or acute, lateral teeth prominent, length at least one-half that of median teeth, margin of head hardly sexdentate ( Fig. 106 View Figures 102–109 , 112 View Figures 110–119 ). Pronotal puncturing always conspicuous (×10; Fig. 105 View Figures 102–109 ). Subhumeral (8 th) elytral stria always at least finely carinulate ( Fig. 107 View Figures 102–109 , 115 View Figures 110–119 ); elytral striae usually uniform, although base of 7 th usually effaced at elytral umbone. Compressed distal portion of parameres (viewed laterally, Fig. 113 View Figures 110–119 ) very short, truncate (“snub nosed”). Southern Arizona and west coast of mainland Mexico....................................................................... 12
12. Dorsum dark metallic green or yellow-green, rarely black; venter with green reflections on legs and pleura ( Fig. 102–103, 107 View Figures 102–109 ). Anterior ends of 2 nd and 3 rd elytral interstriae not swollen or only very weakly so ( Fig. 102–103 View Figures 102–109 ). Western coast of mainland Mexico from far southern Sonora to Nayarit ( Fig. 45 View Figure 45 ).............................................................. Boreocanthon forreri (Bates) View in CoL [p. 56]
— Dorsum and venter black, often with green or blue sheen (intensified when surface is wet). Anterior ends of 2 nd and 3 rd elytral interstriae distinctly swollen ( Fig. 110–111 View Figures 110–119 ). South-central Arizona southward into northwestern Sonora ( Fig. 45 View Figure 45 )......................... Boreocanthon melanus (Robinson) View in CoL [p. 58]










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Boreocanthon Halffter, 1958
| Edmonds, W. D. 2022 |
Canthon
| Halffter G & Martinez A. 1977: 81 |
Boreocanthon
| Halffter G. 1958: 208 |
abrasus
| Howden HF 1966: 732 |
| Robinson M. 1948: 87 |
| Robinson M. 1948: 88 |
| Robinson M. 1948: 87 |
| Robinson M. 1948: 91 |
| Pierce WD 1946: 122 |
| Balthasar V. 1939: 229 |
| Schaeffer C. 1915: 50 |
| Schaeffer C. 1915: 50 |
| Bates HW 1887: 35 |
| Bates HW 1887: 31 |
| Bates HW 1887: 31 |
| Horn GH 1870: 45 |
| Horn GH 1870: 46 |
| Horn GH 1870: 46 |
| Horn GH 1870: 46 |
| Harold 1868: 129 |
| Harold 1868: 68 |
| LeConte JL 1866: 381 |
| LeConte JL 1859: 11 |
| LeConte JL 1859: 11 |
| LeConte JL 1859: 10 |
| LeConte JL 1857: 41 |
| Say T. 1823: 208 |
| Say T. 1823: 208 |