
Nephtys paradoxa Malm, 1874
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2682.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:CC2B98CA-8CEB-4362-A018-031A4B27A725 |
|
persistent identifier |
https://treatment.plazi.org/id/CA4887EA-B745-9B6C-7997-04381AB64EFF |
|
treatment provided by |
Felipe |
|
scientific name |
Nephtys paradoxa Malm, 1874 |
| status |
|
Nephtys paradoxa Malm, 1874 View in CoL
Figure 16 View FIGURE 16 , 23 View FIGURE 23
Nephthys paradoxa Malm, 1874: 78 View in CoL , pl. I, fig. 2; Levinsen 1887; Heinen 1911: 36; Augener 1912: 204; Fauvel 1914: 199; Fauvel 1923: 375, fig. 146F–I; Augener 1928: 701; Ditlevsen 1937: 19; Bellan 1960: 13; Bellan 1961: 265; Foret-Montardo 1969: 818.
Nephthys pansa View in CoL [nomen oblitum] Ehlers, 1874: 293; Ehlers 1875: 40, pl. III, figs. 1 and 2; Horst 1881: 7; McIntosh 1908: 40, figs. 39, 40.
Nephthys phyllobranchia McIntosh, 1885: 164 View in CoL , pl. XXVI, fig. 10; pl. XXVII, fig. 3; pl. XIVA, figs. 12 and 13; Hartman 1950: 111.
Nephthys hombergii Heinen 1911: 16 (partim) (not Savigny in Lamarck, 1818).
? Nephtys schmitti Hartman, 1938: 152 View in CoL , fig. 65; Hilbig 1997: 342, fig. 13.11.
Nephtys paradoxa Hartman 1944: 335 View in CoL , 339, pl. XV, fig. 6; Hartman 1950: 111; Pettibone 1954: 271, fig. 30J–K; Eliason 1962: 249; Fauchald 1963: 13, figs. 1A, 2B and 3C; Pettibone 1963: 200, fig. 47D; Hartman 1965: 92; Hartman 1967: 81; Wolff 1968: 4, fig. 4; Kirkegaard 1969: 55, fig. 19; Bellan 1969: 42; Hartmann-Schröder 1971: 214; Amoureux 1972: 66; Fauchald 1972: 91; Hartmann-Schröder 1974: 207; Paxton 1974: 204;? Rozbaczylo and Castilla, 1974: 201; Rainer and Hutchings 1977: 338, fig. 43; Hartmann-Schröder 1977: 88; Campoy 1982: 513; Imajima and Takeda 1987: 50, figs. 5A–I and 6; Jirkov, 1989: 78, fig. 16.6; Rainer 1991: 81, fig. 3E; Kirkegaard 1992: 341, fig. 167; Kirkegaard 1995: 39; Hartmann-Schröder 1996: 230; Dnestrovskaya and Jirkov 2001: 206, 1 fig; Laborda 2004: 408, fig. 150A–B.
Nephtys brachycephala Uschakov 1955: 216 View in CoL , fig. 69F–G (not Moore 1903).
Nephtys (Nephtys) paradoxa Day 1967: 347 View in CoL , fig. 15.2N –O.
Type locality. Koster , Bohuslän, Sweden .
Material examined. Norway, Trondheimsfjord , Vannviksbukt: RV Harry Borthen, 63º32.745’– 63º32.160’N, 10º14.910’– 10º14.540’E, 200 m, Sneli-sledge, Feb 2003, 1 incomplete spm ( MB36000140 ) GoogleMaps . Sweden, Skagerrak , Bohuslän: 58º24.024’– 58º24.255’N, 10º38.147’– 10º40,005’E, 218–264 m, Aug 2006, 1 complete spm ( MB36000159 ) GoogleMaps ; Koster : 120 fms, Jul 1965, 1 incomplete spm, syntype ( GNHM Polych. 55) ; Kosterfjorden : 120 fms, Jul 1965, 1 incomplete spm, syntype ( GNHM Polych. 1208) . Gulf of Cadiz , Pen Duick Escarpment: TTR12 cruise, RV Prof. Logachev, 35º17.695’N, 6º47.082’W, 560 m, grab, Jul 2002, 1 complete spm ( DBUA 00673-01 View Materials ) GoogleMaps ; TTR16 cruise, RV Prof. Logachev , 35º17.693’N, 6º47.089’W, 556 m, grab, May 2006, 1 complete spm ( MB36000128 ) GoogleMaps . American coast, off Delaware: Challenger Expedition, 38º34’N, 72º10’W, 1240 fms, May 1873, 1 incomplete spm ( NHM 1885.12 .1.128, as N. phyllobranchia , holotype) GoogleMaps ; cruise 9504, RV Albatross IV, 43º38’N, 68º56’W, 126 m, Sep 1995 ,1 incomplete spm ( MCZ 37222) GoogleMaps ; RV Eastward , 33º38’N, 75º51’W, 2300 m, Mar 1972, 1 spm in two fragments ( MCZ 37209) GoogleMaps .
Pacific Ocean. Alaska, NE Gulf of Alaska: 58º32.20’N, 139º32.60’W, 261 m, Nov 1979, 1 complete spm ( CASIZ BLM 36208, as N. schmitti ); South Alaska Peninsula: RV Albatross, 54º00.00’N, 162º40.30’W, 483 fms, 2 complete spms ( USNM 20323 View Materials ) GoogleMaps .
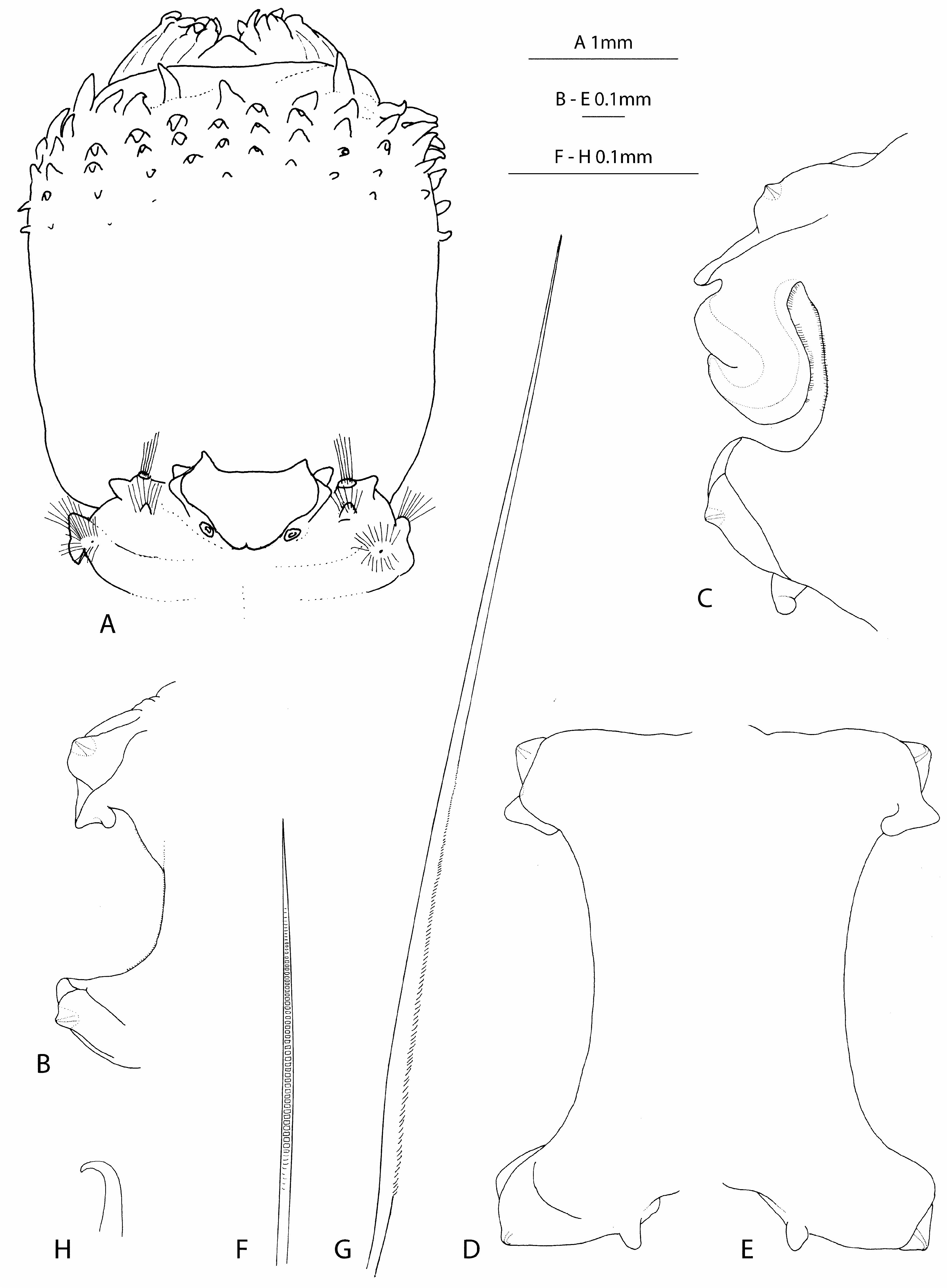
Description. Examined specimens up to 179 mm long for up to 91 chaetigers. See Fig. 16 View FIGURE 16 for length and width measurements. Body stout, slightly wider anteriorly, gradually tapering from middle region to pygidium. Poor dorsal delineation between anterior segments. Colour in ethanol brownish with darker brown pigment on prostomium and anterior segments; some specimens with purple spots on prostomium; chaetae amber; tips of aciculae red. Eyes not visible. Pharynx distal region with 10 pairs of terminal bifid papillae, separated by dorsal and ventral simple, conical papillae (in some specimens the dorsal simple papilla is low and rounded); middorsal papilla slightly larger than subterminal ones ( Fig. 16D View FIGURE 16 ); midventral papillae, if present, similar in size and shape to subterminal ones; subdistal region with 22 rows of (3)4–6 conical subterminal papillae, extending over 1/3 length of pharynx ( Fig. 23A View FIGURE 23 ); proximal region without warts but usually wrinkled. Jaws not examined. Prostomium subquadrangular (subpentagonal when pharynx everted), anterior margin straight, posterior margin U-shaped; antennae and palps conical, very short; palps inserted ventrolaterally on anterior region of prostomium, slightly behind antennae. Nuchal organs conspicuous, rounded. Parapodia biramous; interramal space “U-shaped”; moderately ciliated; posterior parapodia with well separated rami. Parapodia of chaetiger 1 equal in size to subsequent ones, directed anteriorly, parallel to prostomium; notopodial acicular lobes conical, prechaetal lamellae rudimentary, postchaetal lamellae well developed but not extending beyond acicular lobes, rounded; neuropodial pre- and postchaetal lamellae forming a cylinder covering acicular lobe; dorsal cirri small, rounded; ventral cirri small, conical, slightly larger than dorsal cirri ( Fig. 16C View FIGURE 16 ). Acicular lobes of following parapodia rounded with a "conical tip" due to acicula, becoming conical in posterior parapodia; pre- and postchaetal lamellae poorly developed (prechaetal lamellae almost rudimentary), surrounding inner part of acicular lobes in each rami; dorsal cirri small, lamelliform and concave dorsally (as an extension of pre- and postchaetal lamellae); ventral cirri conical, small ( Figs. 23B–E View FIGURE 23 ). Branchiae recurved, with membranous expansions externally and internally when fully developed; moderately ciliated; present from chaetigers 9–14, where it is still very rudimentary, well visible from chaetigers 13–15; become rudimentary again in posterior chaetigers and vestigial or absent in posteriormost chaetigers (from about chaetiger 60–65); occupy all interramal space when fully developed. Chaetae short, of three kinds: barred chaetae in preacicular position ( Fig. 23F View FIGURE 23 ), spinulated chaetae in postacicular position ( Fig. 23G View FIGURE 23 ), and capillary chaetae in neuropodia of chaetiger 1. One acicula per ramus, posterior ones with curved tips ( Fig. 23H View FIGURE 23 ).
Remarks. Fauchald (1963) provided an earlier history of the synonyms of N. paradoxa species. He referred to “ N. phyllocirra McIntosh ” as a possible synonym of N. paradoxa , thus accepting the earlier suggestion by Hartman (1950). However, the species that Hartman suggested to be a possible synonym of N. paradoxa was N. phyllobranchia McIntosh and not N. phyllocirra . Furthermore, the authorship of N. phyllocirra is Ehlers and not McIntosh as mentioned by Fauchald (1963). Assuming these corrections, Fauchald was in fact referring to N. phyllobranchia (and not N. phyllocirra ) as a synonym of N. paradoxa . Nephtys phyllocirra was described by Ehlers (1887) for specimens from off southern Florida and differs mainly by having cirriform branchiae from chaetiger 6 until the end of body.
Imajima and Takeda (1987) synonymized N. schmitti Hartman, 1938 from Alaska to N. paradoxa , although with doubts, since they had not examined the type material. Later, Hilbig (1997) examined the holotype of N. schmitti along with other N. schmitti specimens from California, as well as specimens of N. paradoxa from Alaska, and concluded that the two species differed by the number of pharynx terminal papillae (20 in N. schmitti , 22 in N. paradoxa ), the shape of subterminal papillae (short in N. schmitti , long and slender in N. paradoxa ), and the occurrence of foliaceous branchiae on segments 12–30 in N. schmitti and on segments 20–65 in N. paradoxa . We examined specimens of N. schmitti from Alaska (including the holotype) and specimens of N. paradoxa from Europe (including the type locality) and the eastern coast of USA. Apart from slightly better developed postchaetal lamellae in the specimens of N. schmitti , no consistent differences were found between these specimens and the European ones referred to N. paradoxa . The 22 terminal bifid papillae mentioned by Hilbig (1997) are in fact 20 (10 on each side), separated dorsally and ventrally by a simple papilla, and this was observed in all specimens examined (including the ones from USA). Significant differences in the length of subterminal papillae in the two species were not found, although in the specimens of N. schmitti and the European ones of N. paradoxa the papillae seem to be slightly broader basally. Both N. schmitti and the European specimens of N. paradoxa have foliaceous branchiae between chaetigers 12–42. However, in the specimens of N. paradoxa from USA the branchiae start being foliaceous only in the middle chaetigers (from 25–26 until 40–64), and the postchaetal lamellae are of the same size or slightly larger than the acicular lobes (instead of poorly developed as in the specimens from Europe). Therefore, N. schmitti is here considered to belong to a N. paradoxa species complex. According to the many records from deep-waters from very different geographical regions N. paradoxa seems to have a worldwide distribution. However, many of these records including N. paradoxa from North America should be carefully revised with examination of more specimens. The taxonomical status of this species complex can possibly be resolved only by molecular analyses.
Distribution. Arctic Ocean; Atlantic Ocean ( Greenland; from Iceland to South Africa and from Gulf of St. Lawrence to off Delaware); Mediterranean Sea (SE Spain); Pacific Ocean (Bering Strait, Bering Sea, Sea of Okhotsk, Japan; Australia; off Guatemala; off Peru; off Chile; Magellan Strait) ( Fauchald 1972; Rozbaczylo & Castilla 1974; Imajima & Takeda 1987; Kirkegaard 1995; Laborda 2004).
Habitat. Muddy sediments; 50–8000 m depth ( Rainer & Hutchings 1977; Laborda 2004).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nephtys paradoxa Malm, 1874
| Ravara, Ascensão, Cunha, Marina R. & Pleijel, Fredrik 2010 |
Nephtys (Nephtys) paradoxa
| Day, J. H. 1967: 347 |
Nephtys brachycephala
| Uschakov, P. V. 1955: 216 |
Nephtys paradoxa
| Laborda, A. J. 2004: 408 |
| Hartmann-Schroder, G. 1996: 230 |
| Kirkegaard, J. B. 1995: 39 |
| Kirkegaard, J. B. 1992: 341 |
| Rainer, S. F. 1991: 81 |
| Jirkov, I. A. 1989: 78 |
| Imajima, M. & Takeda, Y. 1987: 50 |
| Campoy, A. 1982: 513 |
| Rainer, S. F. & Hutchings, P. A. 1977: 338 |
| Hartmann-Schroder, G. 1977: 88 |
| Hartmann-Schroder, G. 1974: 207 |
| Paxton, H. 1974: 204 |
| Rozbaczylo, N. & Castilla, J. C. 1974: 201 |
| Amoureux, L. 1972: 66 |
| Fauchald, K. 1972: 91 |
| Hartmann-Schroder, G. 1971: 214 |
| Kirkegaard, J. B. 1969: 55 |
| Bellan, G. 1969: 42 |
| Wolff, W. J. 1968: 4 |
| Hartman, O. 1967: 81 |
| Hartman, O. 1965: 92 |
| Fauchald, K. 1963: 13 |
| Pettibone, M. H. 1963: 200 |
| Eliason, A. 1962: 249 |
| Pettibone, M. H. 1954: 271 |
| Hartman, O. 1950: 111 |
| Hartman, O. 1944: 335 |
Nephtys schmitti
| Hilbig, B. 1997: 342 |
| Hartman, O. 1938: 152 |
Nephthys hombergii
| Heinen, A. 1911: 16 |
Nephthys phyllobranchia
| Hartman, O. 1950: 111 |
| McIntosh, W. C. 1885: 164 |
Nephthys paradoxa
| Foret-Montardo, P. 1969: 818 |
| Bellan, G. 1961: 265 |
| Bellan, G. 1960: 13 |
| Ditlevsen, H. 1937: 19 |
| Augener, H. 1928: 701 |
| Fauvel, P. 1923: 375 |
| Fauvel, P. 1914: 199 |
| Heinen, A. 1911: 36 |
| Malm, A. W. 1874: 78 |
Nephthys pansa
| McIntosh, W. C. 1908: 40 |
| Horst, R. 1881: 7 |
| Ehlers, E. 1875: 40 |
| Ehlers, E. 1874: 293 |