
Pycnoderma dannyi, Salazar-Vallejo, Sergio I., 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.277211 |
|
DOI |
https://doi.org/10.5281/zenodo.6183611 |
|
persistent identifier |
https://treatment.plazi.org/id/D34C87B8-4D12-260C-FF44-FC066276FC15 |
|
treatment provided by |
Plazi |
|
scientific name |
Pycnoderma dannyi |
| status |
sp. nov. |
Pycnoderma dannyi View in CoL n. sp.
Figure 14 View FIGURE 14
Stylarioides congoense: Fauvel, 1939:17 –18 (partim); Kirkegaard, 1959:39 –40 (partim).
Type material. Tropical Eastern Atlantic Ocean. Holotype (ZMUC-1782), off Port Marshall, Liberia, R/V Atlantide Sta. CII-1 (53 in publication), 12 m, 7 Jan. 1946, J.B. Kirkegaard, coll. One paratype (ZMUC-1783), Guinea Bay, off Northwestern Angola, R/V Galathea, Sta. 86 (06°19.3ʹ S, 12°06.5ʹ E), 40 m, 8 Dec. 1950.
Additional material. One specimen ( IRFA), off Kipundji, Congo, 25 m, 26 Aug. 1965, A. Crosnier, coll. (mature female, anterior fragment, broken into two pieces, fixed in alcohol, some parapodia previously removed; chaetigers 2–6 very long, pale, chaetigers 4–5 with anterior margin projected over the previous segment; 3–4 irregular rows of elongate capitate papillae per segment).
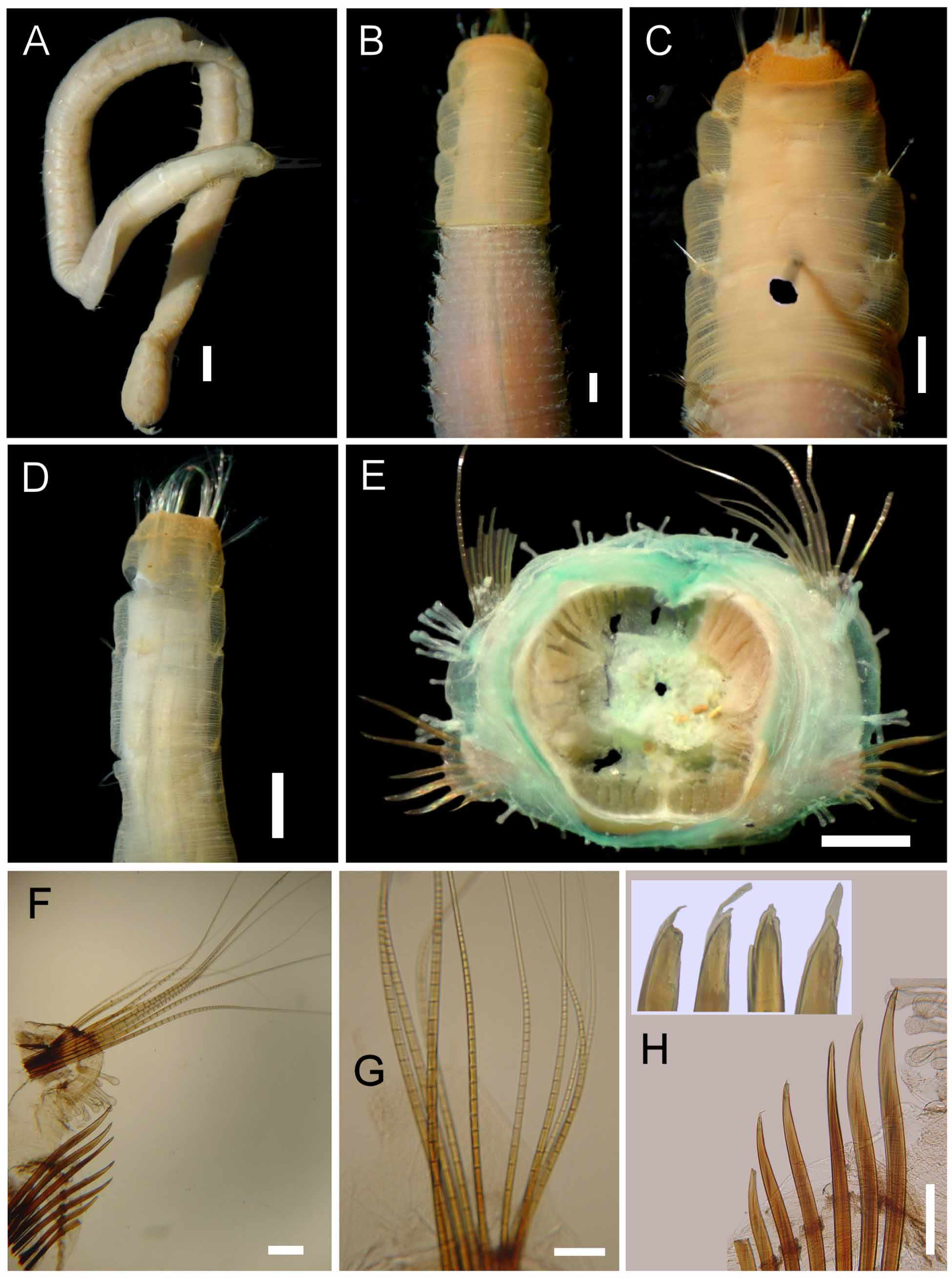
Description. Holotype incomplete, broken into two fragments; body mostly pale-brown, chaetigers 1–6 pale yellow. Body cylindrical ( Fig. 14 View FIGURE 14 A), slightly tapering posteriorly; tunic papillated, free from sediment cover. Papillae reduced in first few chaetigers ( Fig. 14 View FIGURE 14 B–D); most of body with long, capitates papillae, arranged in 3–4 bands per segment ( Fig. 14 View FIGURE 14 E). Holotype 54 mm long, 5 mm wide, cephalic cage 4 mm long, 70 chaetigers.
Anterior end not exposed; not dissected to avoid damage. Cephalic cage chaetae as long as 4/5 body width. Chaetiger 1 involved in the cephalic cage; chaetiger 2 with chaetae longer than following chaetigers, but not contributing to cage; chaetiger 1 with chaetae arranged in short dorsolateral row, neurochaetal lobe almost fused to notochaetal lobe; about 12 notochaetae and eight neurochaetae per bundle.
Anterior dorsal margin of chaetiger 1 finely papillated, not projecting anteriorly ( Fig. 14 View FIGURE 14 B, D). Chaetigers 1–6 mostly smooth, without long papillae; two tiny papillae in transverse depressions slightly ahead of, and behind notopodial lobes. Chaetigers 1–3 becoming progressively longer up to chaetiger 6; chaetiger 7 shortest, then following segments of similar length, wider than long, more papillose. Chaetal transition from cephalic cage to body chaetae abrupt; neurohooks start in chaetiger 8. Gonopodial lobes not seen ( Fig. 14 View FIGURE 14 C).
Parapodia well developed, lateral ( Fig. 14 View FIGURE 14 E, F); median neuropodia ventrolateral. Notopodia low, rounded lobes without suprachaetal papillae; infrachaetal papillae very long, capitate, 4–6 in a row, directed ventrally (6–8 in posterior chaetigers). Neuropodia low, rounded lobes with 4–5 slightly smaller suprachaetal papillae (2–4 in posterior chaetigers), and 3–4 infrachaetal papillae (1–2 in posterior chaetigers), smaller than the superior ones.
Median notochaetae long, arranged in short transverse row, 6–8 notochaetae per bundle, as long as 1/2 body width; notochaetae of chaetigers 1–6 multiarticulated capillaries, articles short medially, shorter distally; other notochaetae multiarticulated capillaries with long articles basally and medially, distally hyaline, articles short along large chaetal proportion ( Fig. 14 View FIGURE 14 G). Neurochaetae multiarticulated capillaries in chaetigers 1–6; falcate neurohooks from chaetiger 7, arranged in transverse row, anterior chaetigers with five per fascicle, up to seven in posterior chaetigers. Neurohooks brown, oligo-articulate, aristate, most eroded or broken distally ( Fig. 14 View FIGURE 14 H); articles short basally, become longer medially, distally very long.
Posterior end unknown.
Etymology. This species is named after Danny Eibye-Jacobsen for his many contributions to polychaete systematics, and especially because of his interest in promoting the study of polychaetes in his country and abroad. The latter has resulted in being involved as editor and author of an impressive contribution to our knowledge of Northeastern Indian Ocean polychaetes.
Type locality. off Port Marshall, Liberia, 12 m depth.
Variation. The paratype is an anterior fragment, 35.5 mm long, 1.5 mm wide, cephalic cage 3 mm long, 43 chaetigers; first neurohooks from chaetiger 7.
Remarks. Pycnoderma dannyi n. sp. is closely allied to P. congoense ; the two species differ because P. dannyi n. sp. has longer anterior chaetigers, an earlier start of the neurohooks, and fewer rows of large, capitate papillae along the body. Fauvel (1939) noticed some differences between his specimen and the published accounts of P. congoense , especially concerning the type of neurochaetae; he found some neurohooks in his specimen. Further, he noticed that the neurohooks began on chaetiger 7, and that there were larger papillae associated with the chaetal bundles. These observations were confirmed by Kirkegaard (1959). Consequently, their materials are herein regarded as a distinct species, and these features are regarded as diagnostic for P. dannyi .
Distribution. Western Tropical Africa, from Gambia to Northwestern Angola, in shallow water (12–40 m).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pycnoderma dannyi
| Salazar-Vallejo, Sergio I. 2011 |
Stylarioides congoense:
| Kirkegaard 1959: 39 |
| Fauvel 1939: 17 |