
Polydora hoplura Claparède, 1868
|
publication ID |
https://doi.org/10.11646/zootaxa.4282.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:A896C27D-2063-4CCD-8E69-B788DC337FD3 |
|
DOI |
https://doi.org/10.5281/zenodo.6033520 |
|
persistent identifier |
https://treatment.plazi.org/id/D522E54A-7214-FFDC-FF12-FDB8FEC365A2 |
|
treatment provided by |
Plazi |
|
scientific name |
Polydora hoplura Claparède, 1868 |
| status |
|
Polydora hoplura Claparède, 1868 View in CoL
( Figs 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Polydora hoplura Claparède, 1868: 318 View in CoL –319, pl. XXII, fig. 2; 1869: 58–59, pl. XXII, fig. 2; 1870: 58–59, pl. XXII, fig. 2. Marion & Bobretzky 1875: 84. Carazzi 1893: 20 –21, pl. 2, figs 6, 7, 13, 16, 18. Lo Bianco 1893: 30; 1909: 584. Saint- Joseph 1894: 65. Soulier 1903: 83 –86, fig. 12. Douvillé 1907: 364 –365, fig. 7. McIntosh 1909: 173 –174; 1915a: 212–213; 1915b: pl. 101: fig. 10, pl. 106: fig. 6; 1923: 486–487. Fauvel 1927: 50, fig. 17a–g. Wilson 1928: 578 –585, textfig. 2, pls V–VII (larval morphology). Read 1975: 411 –412, fig. 6. Blake & Kudenov 1978: 264, fig. 47. Hutchings & Turvey 1984: 15. Colognola et al. 1984: 748. Lleonart 2001: figs 4–6, 9, 26–32. David et al. 2014: figs 4–6 (larval morphology). Radashevsky & Migotto 2016: 2 –7, figs 2–5 (adult and larval morphology). Sato-Okoshi et al. 2016: 3 –6, figs 6, 7.
Polydora ( Polydora) hoplura: Rioja 1931 ( Part.) View in CoL : 70, pl. 19, figs 8–13. Hartmann-Schröder 1971: 305; 1996: 318.
Polydora hoplura hoplura: Day 1967: 468 View in CoL , fig. 18.2k–m.
Leucodora sanguinea Giard, 1881: 71 –73. Fide Dollfus 1921: 17; 1932: 275.
Polydora uncinata Sato-Okoshi, 1998 View in CoL : pp. 278–280, fig. 1; 1999: p. 835. Radashevsky & Olivares 2005: 491 –494, figs 2–4. Sato-Okoshi et al. 2008: 493 –495, figs 2–3; 2012: 87, figs 4A–B, D. Sato-Okoshi & Abe 2012: 43 –44, fig. 3. Fide Radashevsky & Migotto 2016: 2; Sato-Okoshi et al. 2016: 4.
Material. Italy, Tyrrhenian Sea, Campania, Gulf of Naples, Port of Ischia , Ischia Is., 40.74439°N, 13.93948°E, intertidal, Radashevsky, V.I. & M.C. Gambi, 0 7 Jul 2014: from shell of live whelk Stramonita haemastoma (Linnaeus, 1767) , SMF 24359 ( neotype), two individuals from shells of live whelk S. haemastoma and two individuals from shell of live European flat oyster Ostrea edulis Linnaeus, 1758 GoogleMaps , MIMB 28148 View Materials (4). Tyrrhenian Sea, Lazio, Fiumicino , 41.7739°N, 12.2189°E, from a shell in fouling of artificial platform, st. 5, 5 m, spring 1979, Gambi, M.C. GoogleMaps , MIMB 33 View Materials 0 27 (34), 33028 (7). Ionian Sea, Apulia, Taranto , 40.4323°N, 17.2409°E, 1 m, from shells of the oyster O. edulis, Radashevsky, V.I. , 15 Jul 2014 GoogleMaps , MIMB 33029 (23).
France, Brittany, La Manche, NE to Roscoff , 48.7487°N, 3.9062°W, 40 m, from empty scallop shell, Radashevsky, V.I. & C. Houbin, 27 May 2014, MIMB 3 View Materials 3030 (1) GoogleMaps . Aquitaine, Bay of Biscay, Arcachon Bay: Jacquets , 44.7212°N, 1.1887°W, from shells of Pacific oyster Crassostrea gigas (Thunberg, 1793) , Radashevsky, V.I. & S. Gasmi, 12 Jun 2014, MIMB 33031 View Materials (22); la Chapelle, 44.6645°N, 1.1802°W, 9 m, from shells of Pacific oyster C. gigas, Radashevsky, V.I. & B. Gouillieux , 13 Jun 2014, MIMB 33032 View Materials (25). Legallais, Arcachon, 44.663914°N, 1.175977°W, rocky intertidal, from shell of gastropod Ocenebra erinaceus (Linnaeus, 1758) occupied by hermit crab Clibanarius erythropus (Latreille, 1818) , Lavesque, N., 16 Nov 2016, MIMB 33064 View Materials (3) GoogleMaps . Languedoc-Roussillon, Gulf of Lion, Leucate , 42.8778°N, 3.0229°E, 2 m, from shells of the Pacific oyster C. gigas, Radashevsky, V.I. & C. Labrune , 26 Jun 2014, MIMB 33033 View Materials (19). GoogleMaps
South Korea, East China Sea , Jeollanam-do Province, Wando County, from shells of abalone Haliotis discus hannai Ino, 1953 cultivated in cages: Sinji Is., 34.345°N, 126.886°E GoogleMaps , 30 Mar 2005, MIMB 33034 (7); Bogil Is., 34.1825°N, 126.5314°E, 7 May 2013, MIMB 33035 (8); Keumil Is., 34.4611°N, 127.0275°E, 27 May 2013, MIMB 33036 (30); Nohwa Is., 34.226117°N, 126.42 5833°E, 0 7 May 2013, MIMB 3 3037 (17); Seohwa Is., 34.317°N, 126.5075°E, 20 Nov 2013, MIMB 33038 (17); 34.191°N, 126.635°E, 27 Nov 2013, MIMB 33039 (13); Soan Is., 34.191°N, 126.635°E, 14 Jun 2016, MIMB 33 View Materials 0 57 (8). Yeocheon County, Yeosu , 34.6344°N, 127.6408°E, from shells of abalone cultivated in land-based tanks GoogleMaps , 12 Nov 2013, MIMB 33 View Materials 0 40 (2). Gyeongsangnam-do Province, Geoje Is., from shells of cultivated Pacific oyster C. gigas : 34.82°N, 128.5°E GoogleMaps , 22 Oct 2013, MIMB 33041 (4); 34.97°N, 128.62°E, 31 May 2016, MIMB 33 0 56 (7). Jeju Province, Jeju Is., from shells of abalone H. discus hannai cultivated in land-based tanks: 33.472252°N, 126.912332°E GoogleMaps , 23 May 2013, KIOST BSMA 2 (15), MIMB 28076 View Materials (120+). Cheongjeonghae Fisheries , 33.35 6012°N, 126.182154°E , 28 Aug 2013, MIMB 33 View Materials 0 42 (13). Dongjin Fisheries , 33.435425°N, 126.266602°E GoogleMaps , 28 Nov 2013, KIOST BSMA 12 (7), MIMB 3 3043 (12); 28 Mar 2014, MIMB 33048 (20+); 30 Apr 2 0 14, MIMB 33049 (10+); 30 May 2014, MIMB 33050 View Materials (20+). Halla Abalone , 33.470857°N, 126.917034°E GoogleMaps , 28 Nov 2013, KIOST BSMA 13 (16), MIMB 33044 (15); 27 Mar 2014, MIMB 33047 View Materials (6+). Jaeil Fisheries , 33.469882°N, 126.918601°E GoogleMaps , 28 Nov 2013, KIOST BSMA 14 (16), MIMB 33045 (100+); 3 Jun 2014, KIOST BSMA 6952 (50+), MIMB 33 0 46 (100+).
Adult morphology (Material from the type locality, Gulf of Naples, Italy). Neotype (SMF 24359) largest complete individual 30 mm long, 1.5 mm wide with 175 chaetigers; other individuals (MIMB 28148) 8 to 11 mm long, with 60 to 100 chaetigers. Pigmentation absent on body. No pigment on palps in a 60-chaetiger individual, and in an individual 11 mm long with 100 chaetigers; 75-chaetiger individual and neotype with five and seven paired black bands on each palp, respectively. Prostomium anteriorly incised, extending posteriorly to end of chaetiger 3 as a low caruncle. Occipital antenna absent in 60- and 75-chaetiger individuals; larger individuals, including neotype, with short occipital antenna on prostomium. Eyes absent in neotype; up to four eyes present or eyes absent in other individuals. Chaetiger 1 with short capillaries only in neuropodia. Chaetiger 5 with up to four dorsal superior winged capillaries, six heavy falcate spines alternating with bilimbate-tipped companion chaetae, and six ventral winged capillaries; falcate spines with lateral flange. Hooded hooks in neuropodia from chaetiger 7, up to ten in a series, not accompanied by capillaries; hooks bidentate, with constriction on shaft. Posterior notopodia (from chaetiger 155 onwards in a 175-chaetiger neotype) each with 1–2 heavy recurved spines and a tuft of slender capillaries; spines with curved tips directed medially. Branchiae from chaetiger 7 continuing posteriorly along most of body (on chaetigers 7–154 in neotype). Pygidium white, cup-shaped with dorsal gap. Of five individuals, two were females (175-chaetiger neotype with oocytes in chaetigers 40–113), two had no gametes, and one was male ( 10 mm long with 75 chaetigers, with branchiae on chaetigers 7–39, and nototrochs composed of single rows of cilia) with spermatocytes, tetrads of spermatids and individual spermatozoa (introsperm with elongated head and midpiece, similar to those in other Polydora species) floating in coelom in chaetigers 29–44.
Other material from Italy and France. Large worms of a size range similar to specimens from type locality (see above). Black pigment diffused on anterior part of prostomium (in front of eyes) in some individuals, varying from weak only on lateral sides to intense on lateral and dorsal sides of prostomium. Similar pigment of variable intensity diffused on dorso-lateral sides of peristomium in some individuals. Up to ten black paired bands present on each palp and black pigment diffused on pygidium in some individuals. Notopodia of 5–16 posterior chaetigers with heavy recurved spines in addition to capillaries. Pygidium flaring disc with wide dorsal gap.
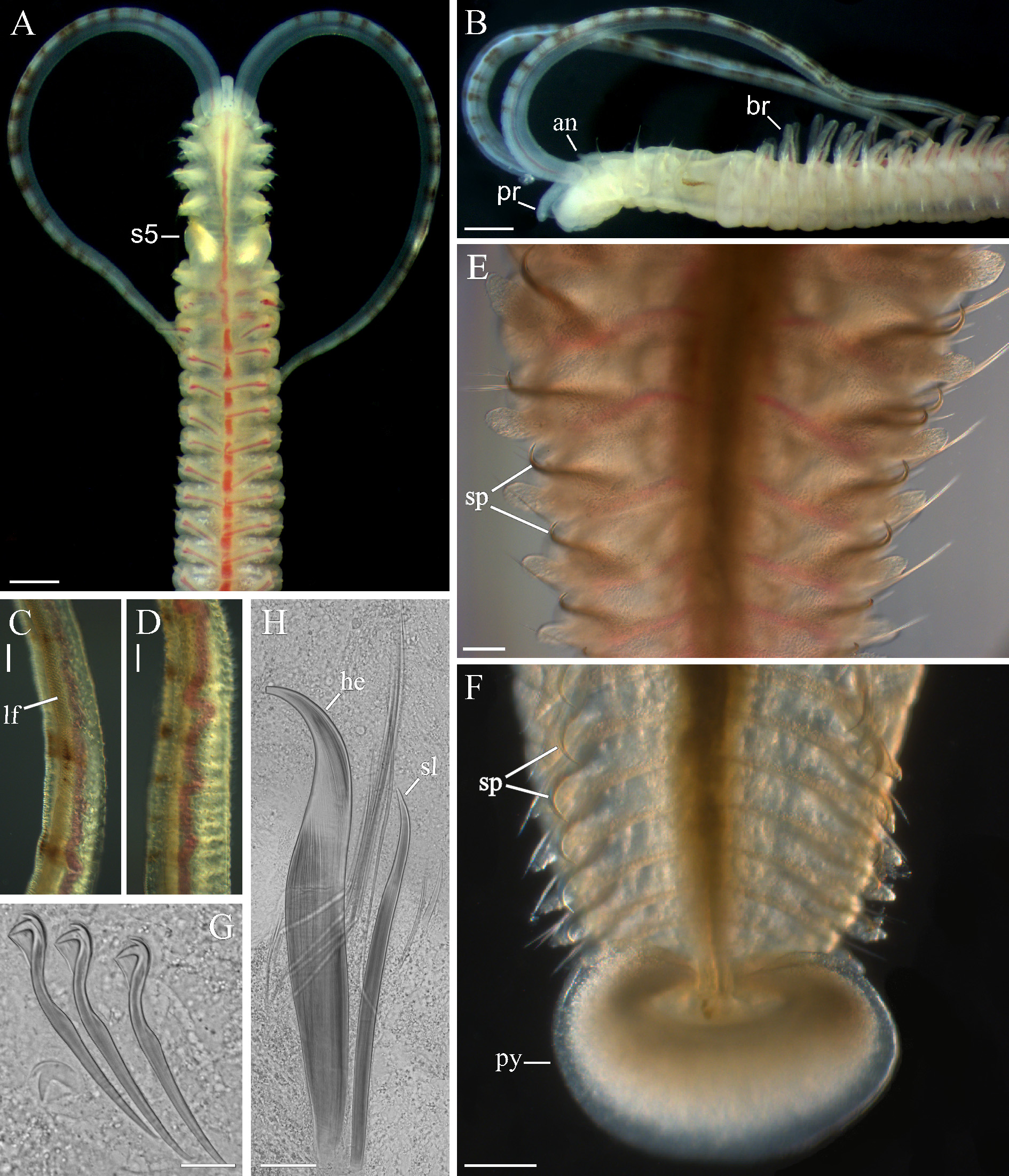
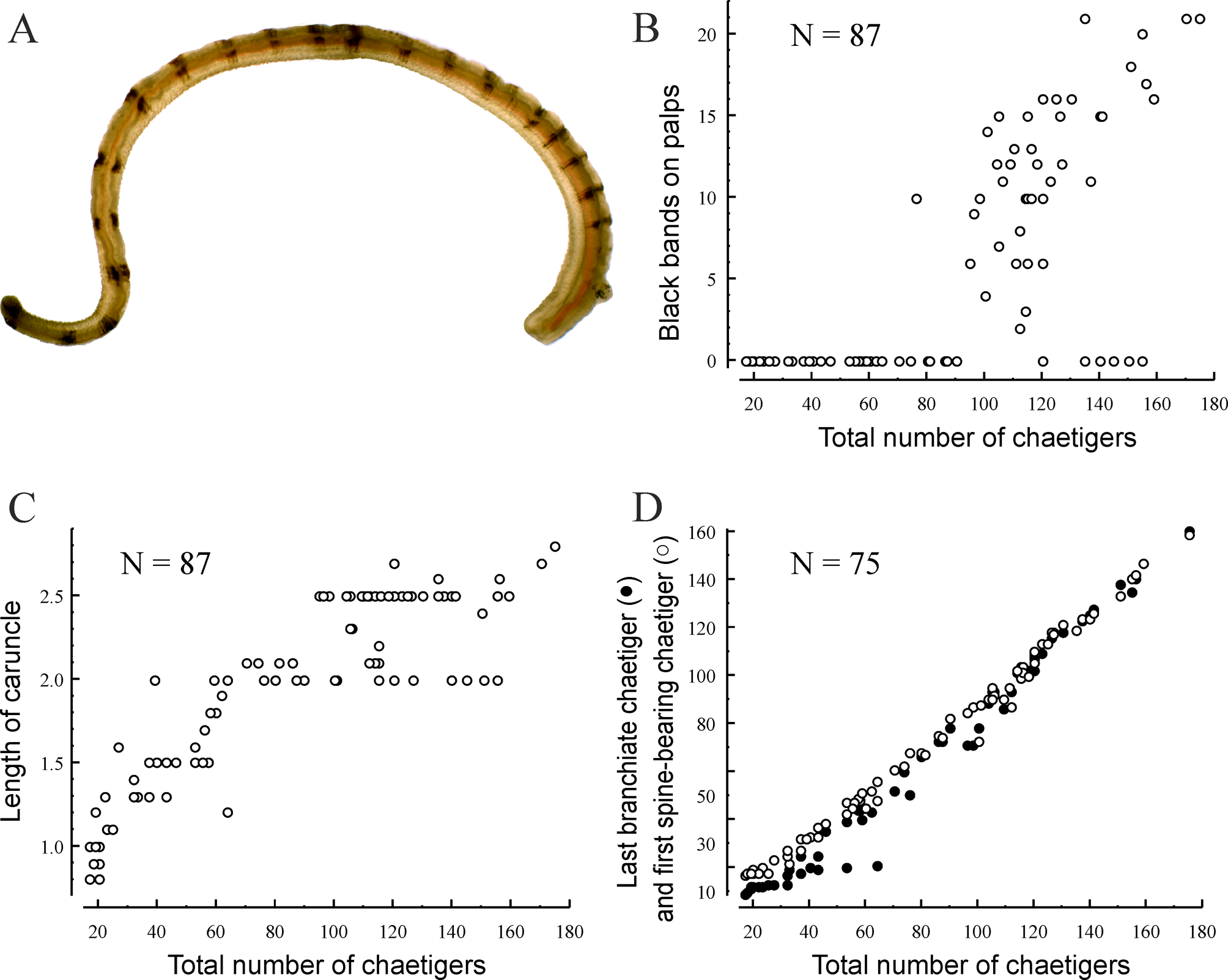
Material from South Korea ( Polydora hoplura in Korean galgori-kin-eolgool-gat-ji-rung-i, meaning a long face worm with hook). Up to 55 mm long and 2 mm wide for 180 chaetigers. Body pale to light tan in life; with numerous glandular cells on dorsal side from chaetigers 10–15 giving dorsum whitish appearance ( Fig. 2 View FIGURE 2 A). Remains of larval melanophores present on dorso-lateral sides of 15–16 anterior chaetigers in individuals up to 50– 60-chaetiger stage; larger worms usually lacking black pigment on body. Black narrow longitudinal stripes on lateral sides of anterior part of prostomium and a pair of small black patches on dorsal side of peristomium present in three individuals (of more than one thousand examined). Up to 25 black paired bands present on each palp in individuals with more than 50 chaetigers ( Fig. 2 View FIGURE 2 A–D); bands usually regularly arranged and of equal intensity along palp ( Fig. 3 View FIGURE 3 A); in some individuals bands more intense and distinct on distal half of palp. Number of bands weakly correlated with total number of chaetigers in an individual ( Fig. 3 View FIGURE 3 B).
Prostomium with shallow anterior incision often seen only in ventral view, posteriorly extending to end of chaetiger 3 (usually to middle of chaetiger 3) as a low caruncle, shorter in small individuals ( Fig. 3 View FIGURE 3 C). Short occipital antenna present on caruncle in individuals with more than 90 chaetigers; smaller individuals usually without antenna. Two pairs of black eyes usually present. Palps as long as 15–30 chaetigers, with longitudinal frontal groove lined with fine cilia, latero-frontal motile compound cilia, and papillae with short non-motile cilia densely arranged in 4–5 rows along both sides of frontal groove ( Fig. 2 View FIGURE 2 C), and also sparsely scattered on lateral and abfrontal palp surfaces all along palp length ( Fig. 2 View FIGURE 2 D).
Chaetiger 1 with short capillaries in neuropodia and small postchaetal lamellae in both rami; notochaetae absent. Notopodia of posterior chaetigers with two kinds of heavy spines in addition to 2–6 slender capillaries ( Fig. 2 View FIGURE 2 H). One spine slender and slightly curved, awl-like ( Fig. 2 View FIGURE 2 H, sl), second spine heavily recurved, sickle-shaped ( Fig. 2 View FIGURE 2 H, he); spines with curved tips directed medially. Slender awl-like spines first appearing in chaetiger 18 in juveniles with 18–19 chaetigers, while heavy sickle-shaped spines first appearing in notopodia in juveniles with 25–30 chaetigers ( Table 1: 25-chaetiger individual). Both kinds of spines beginning from more posterior chaetigers in larger individuals ( Fig. 3 View FIGURE 3 D) what can only be explained by their falling out from anterior notopodia with growth of worms. Slender awl-like spines and heavy sickle-shaped spines developing and falling out at different rates and, therefore, arranged in notopodia differently ( Table 1). In notopodia of new segments developing in growth zone in front of pygidium in worms with more than 25 chaetigers, capillaries and heavy sickle-shaped spines developing first while slender awl-like spines developing later. Consequently, notopodia of a few posteriormost chaetigers bearing only capillaries and heavy sickle-shaped spines. With growth, heavy sickle-shaped spines of anteriormost spine-bearing notopodia falling out first while slender awl-like spines falling out gradually afterwards. Consequently, notopodia of a few anteriormost chaetigers bearing only capillaries and slender awl-like spines. Middle spine-bearing notopodia having both kinds of spines in addition to capillaries. Anterior position of spines correlated with arrangement of branchiae ( Table 1). In small individuals, spines first present from 4–17 chaetigers after last branchiate chaetiger. In mid-size individuals, slender awl-like spines usually from last branchiate chaetiger and heavy sickle-shaped spines usually beginning 1–3 chaetigers after last branchiate chaetiger. In large individuals, slender awl-like spines usually from 1–3 chaetigers before last branchiate chaetiger, while heavy sickle-shaped spines usually beginning in last branchiate chaetiger or in next chaetiger after it.
Chaetiger 5 twice as large as chaetigers 4 and 6, with up to six dorsal superior winged capillaries, seven heavy falcate spines arranged in a slightly curved diagonal row and alternating with bilimbate-tipped companion chaetae, and seven ventral winged capillaries. Dorsal superior and ventral capillaries shorter and fewer than those on adjacent chaetigers. Falcate spines with large subdistal flange on lateral side; upper and inner parts of flange thinner than lower and outer parts, thus in newly developed spines (situated in posterior part of spine row) whole structure appearing as a large tooth joined to main fang by thin sheath; in older spines (situated in anterior part of spine row) lateral flange greatly worn and indistinct.
Hooks in neuropodia from chaetiger 7, up to 12 in a series, not accompanied by capillaries. Hooks bidentate; shaft slightly curved, with weak constriction in upper part ( Fig. 2 View FIGURE 2 G).
Branchiae from chaetiger 7, full-sized from chaetigers 10–12, gradually diminishing in size along posterior half of body, absent on 5–20 posteriormost chaetigers ( Fig. 3 View FIGURE 3 D). Branchiae flattened, with surfaces oriented parallel to body axis, free from notopodial postchaetal lamellae, with longitudinal row of cilia along inner surface.
Nototrochs from chaetigers 7–8 onwards, each composed of one row of short cilia, on branchiate chaetigers extending onto branchiae.
Pygidium disk-like to cup-shaped, usually with distinct dorsal gap, white due to numerous striated glandular cells ( Fig. 2 View FIGURE 2 F).
Glandular pouches in neuropodia from chaetiger 6 in individuals with up to about 30 chaetigers, from chaetiger 7 in larger individuals; pouches large in chaetigers 7–9 and considerably smaller in successive chaetigers.
Digestive tract without ventral buccal bulb and gizzard-like structure.
Nephridia from chaetiger 7 onwards, greenish in life; pairs of nephridia on each chaetiger opening to exterior middorsally via two closely situated nephridiopores.
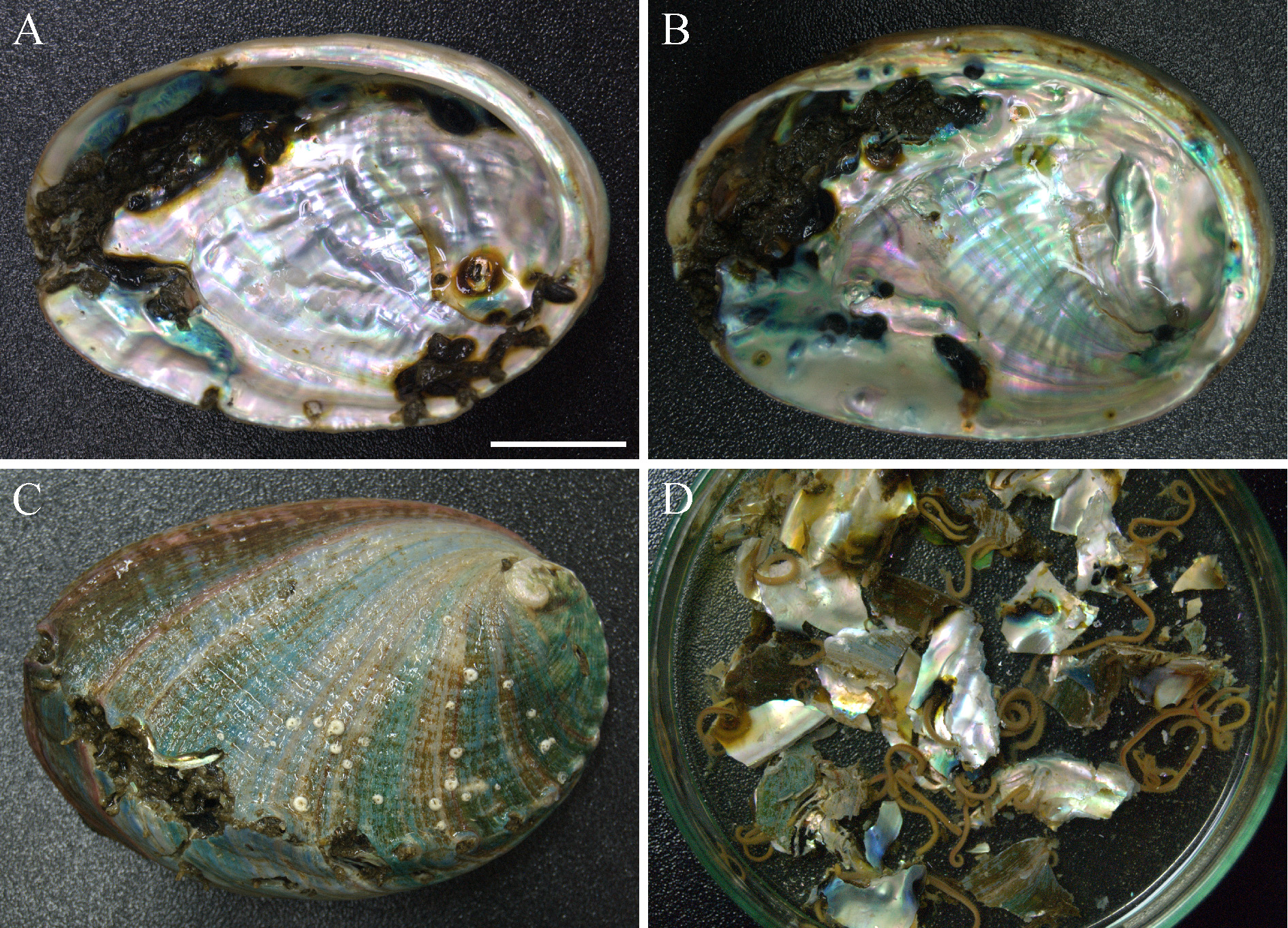
Habitat. Adults of P. hoplura make U-shaped burrows in shells of barnacles, various gastropods and bivalves, including abalone and oysters. Worm burrows appear as detrital tubes inside shells: the walls of the burrows are lined with detritus, and median space of each burrow is also filled with detritus forming a medial wall. Each burrow opens to the outside via two joined apertures forming a characteristic 8-shaped hollow in a shell, and extended by two smooth silty tubes each up to 5 mm long.
In Italy and France, adults of P. hoplura were found in shells of live whelk Stramonita haemastoma , European flat oyster Ostrea edulis , Pacific oyster Crassostrea gigas , empty shells of pilgrim scallop Pecten maximus (Linnaeus, 1758) , and empty shell of gastropod Ocenebra erinaceus occupied by hermit crab Clibanarius erythropus . The worms were rare and did not cause serious damage to the mollusks.
25-chaetiger individual 50-chaetiger individual
chaetiger branchia awl-like sickle-like chaetiger branchia awl-like sickle-like spine spine spine spine 17 42 + In South Korea, P. uncinata (= P. hoplura ) was first recorded in 2004 from the oysters cultivated in Geoje and Goseong (south-eastern part of South Korea), but not found in abalone cultivated in sea cages (Sato-Okoshi et al. 2012). In the present study, we found up to three worms per shell in abalone cultivated in cages in the sea in Wando county, and a maximum of 80 large worms in one shell of abalone cultivated in land-based tanks on Jeju Island ( Fig. 4 View FIGURE 4 D). In many cases worms caused formation of dark brown muddy or nacreous, parchment-like blisters on the inner shell surface ( Fig. 4 View FIGURE 4 A–C). Heavy infestation by P. hoplura occasionally caused secondary bacterial infestation, clogging of respiratory pores and death of mollusk. In contrast to abalone, shells of the Pacific oyster C. gigas widely cultivated around Geoje Island were rarely infested by P. hoplura which seems to not effect oyster condition.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Polydora hoplura Claparède, 1868
| Radashevsky, Vasily I., Choi, Jin-Woo & Gambi, Maria Cristina 2017 |
Polydora uncinata
| Radashevsky 2016: 2 |
| Sato-Okoshi 2016: 4 |
| Sato-Okoshi 2012: 43 |
| Sato-Okoshi 2008: 493 |
| Radashevsky 2005: 491 |
Polydora ( Polydora ) hoplura : Rioja 1931 ( Part .)
| Hartmann-Schroder 1971: 305 |
Polydora hoplura hoplura :
| Day 1967: 468 |
Leucodora sanguinea
| Dollfus 1921: 17 |
| Giard 1881: 71 |
Polydora hoplura Claparède, 1868 : 318
| Radashevsky 2016: 2 |
| Sato-Okoshi 2016: 3 |
| Hutchings 1984: 15 |
| Blake 1978: 264 |
| Read 1975: 411 |
| Wilson 1928: 578 |
| Fauvel 1927: 50 |
| McIntosh 1909: 173 |
| Douville 1907: 364 |
| Soulier 1903: 83 |
| Joseph 1894: 65 |
| Carazzi 1893: 20 |
| Lo 1893: 30 |
| Marion 1875: 84 |
| Claparede 1868: 318 |