
Listriolobus brevirostris Chen and Yeh, 1958
|
publication ID |
https://doi.org/10.12782/specdiv.24.151 |
|
persistent identifier |
https://treatment.plazi.org/id/DB2B5B32-FFAF-A56A-FF16-FB0BFC4606FB |
|
treatment provided by |
Felipe |
|
scientific name |
Listriolobus brevirostris Chen and Yeh, 1958 |
| status |
|
Listriolobus brevirostris Chen and Yeh, 1958 View in CoL (New Japanese Name: Minato Tatejima Yumushi) ( Figs 1–5 View Fig View Fig View Fig View Fig View Fig ; Table 1)
Listriolobus brevirostris Chen and Yeh, 1958: 269–270 View in CoL , 276–278, text-figs 5–7; Stephen and Edmonds 1972: 424; DattaGupta and Menon 1966: 193–195, text-fig. 1A, fig. 2A; Edmonds 1987: 131–132, text-figs 12–13, 22; Li et al. 1994: 206; Wang et al. 1995: 31; Halder 2004: 253; Zhou et al. 2007: 147–149, text-fig. 85; Al-Yamani et al. 2012: 114, pl. 77.
Listriolobus bulbocaudatus Edmonds, 1963: 243–245 View in CoL , pl. 1, fig. 1; Stephen and Edmonds 1972: 423.
Listriolobus aff. bulbocaudatus: Al-Yamani et al. 2009: 25 View in CoL .
Material Examined. Specimens serially dredged in northeastern Osaka Bay in 1999, 6.4–15.2 m deep, fixed in formalin or Bouin’s fluid (abbreviated as B-fixed), collected by I . A. (station data given in Table 1). Registered as NSMT- Ec : 164, 3 June, Stn 7, 1 ind .; 165, 18 August, Stns 7 and 9, 8 inds; 166, 2 Sept ., Stn 7, 2 inds .; 167, 2 September , Stn 9, 1 ind .; 168, 2 September , Stn 34, 1 ind .; 169, 13 September, Stn 34, 2 inds; 170, 27 September, Stn 37, 1 ind .; 171, 13 Oct., Stn 7, 3 inds; 172, 13 October, Stn 9, 1 ind .; 173, 13 October , Stn 26, 1 ind .; 174, 13 October , Stn 34, 1 ind ., B-fixed; 175, 13 October , Stn 37, 1 ind ., B-fixed; 176, 13 October, Stn 37, 3 inds; 177, 26 October, Stn 7, 2 inds; 178, 26 October, Stn 9, 21 inds; 179, 26 October, Stn 37, 2 inds; 180, 23 December, Stn 7, 4 inds, B-fixed; 181, 23 December, Stn 34, 2 inds, B-fixed; 182, 23 December, Stn 37, 1 ind., B-fixed; 183, 23 December , Stn 41, 1 ind ., B-fixed; 184, Near “Kobe Port Island”, Kobe , Osaka Bay, 28 October 1999, I . A. coll., 2 inds; 185, Off “Kobe Port Island”, Kobe, Osaka Bay , 17.5 m deep, 4 November 1999, M . Otani coll ., 1 ind.; 186, Off Kobe, Osaka Bay , 26 November 2001, M . Otani coll ., 1 ind. OMNH-Iv 1820, Off “Kobe Port Island”, Kobe , Osaka Bay, 19 December 2000, M . Hanaoka coll ., 48 inds; 4488, Off Sakai , Osaka Bay, 1 March 2000, H . Ariyama coll ., 6 inds.
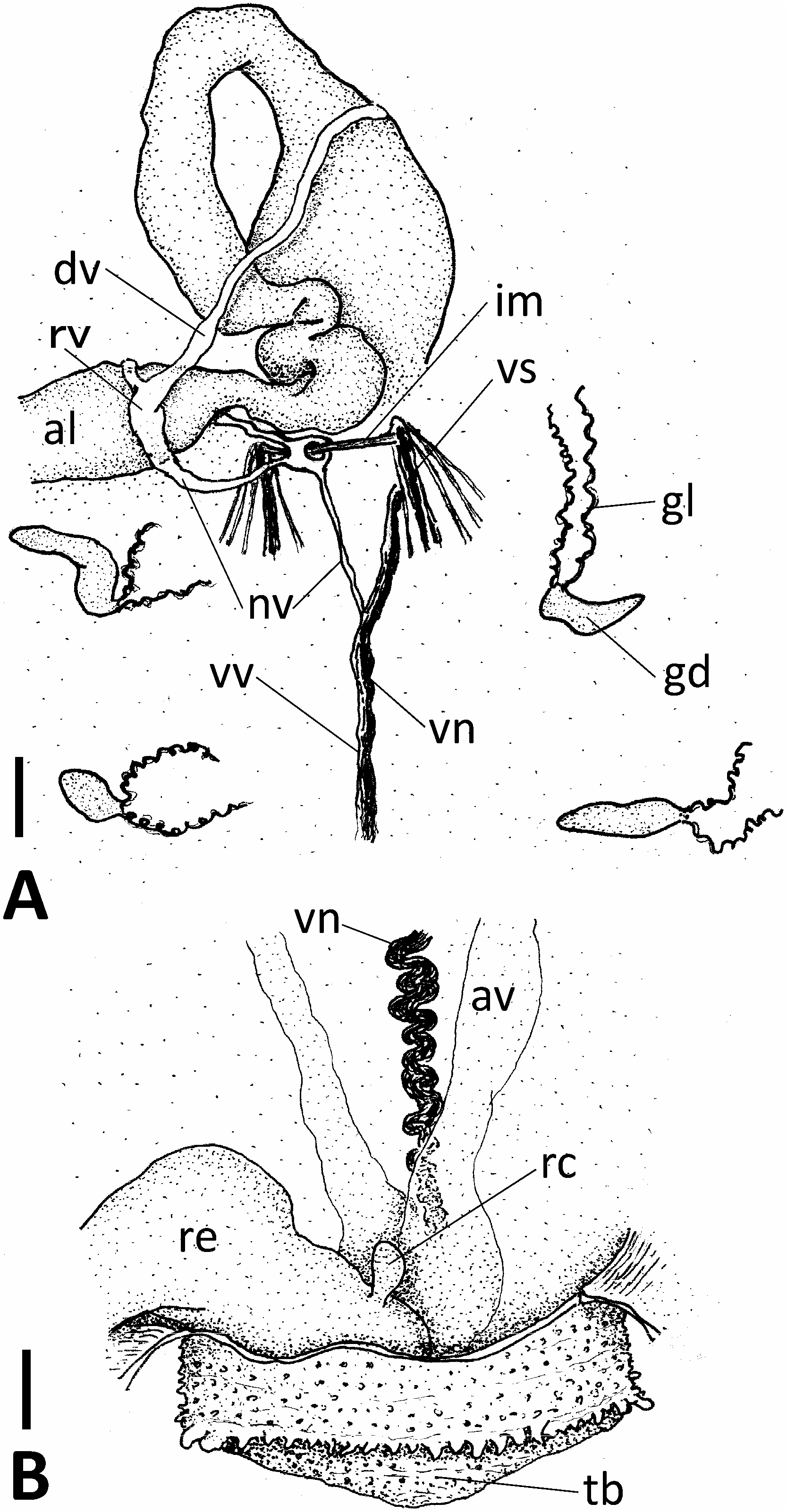
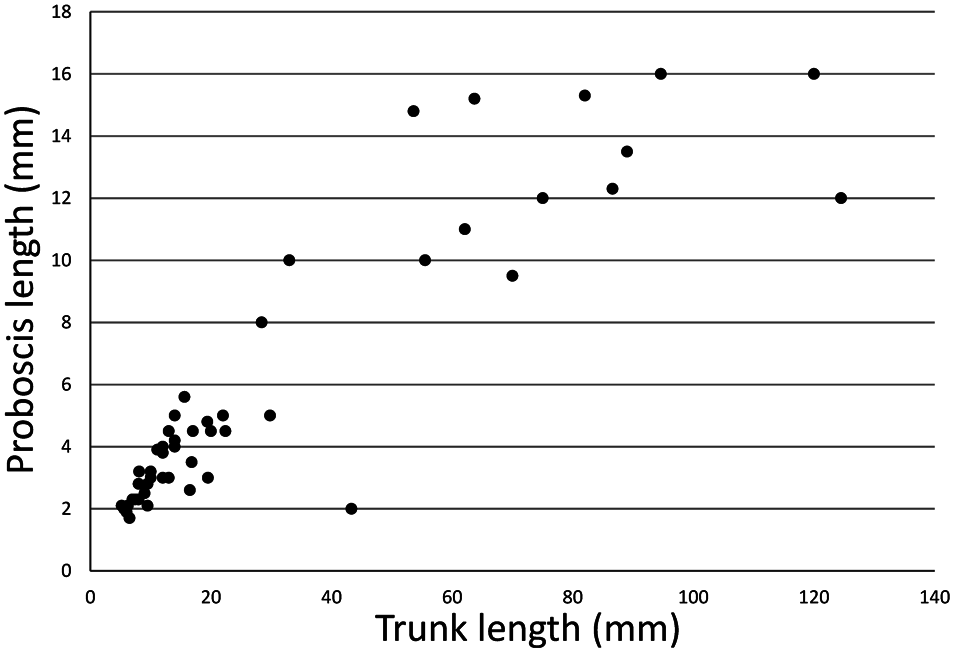
Description. In larger living specimens, trunk predominantly light or dark reddish with narrow white longitudinal lines representing muscle bands, posterior end usually opaque and white due to thick fleshy wall forming a conical terminal bulb; proboscis white to pale yellow, with nearly smooth deeper yellow edges. Reddish trunk coloration and white terminal bulb inconspicuous in smaller specimens. Proboscis 1.7–16.0 mm long, ranging approx. 4–28% (20% in average) of total body length (7.3–136.5 mm) in 52 fixed complete specimens (proboscis/trunk length proportion variable, see Fig. 4 View Fig ); edges separated completely to mouth level, weakly or deeply undulating due to shrinkage at fixation. Trunk cylindrical, up to 124.5 mm long; its surface densely covered with round (sometimes conical at anterior extremity) papillae, up to 0.2 mm high, mostly arranged annularly. Anterior end of terminal bulb fringed with a prominent ring of wart-like papillae, sometimes up to 0.3 mm high ( Fig. 3B View Fig ). Seven longitudinal muscle bands. One pair of ventral setae, their distal one-fourth sickle-shaped; interbasal muscle present. Two pairs of gonoducts posterior to ventral setae; gonostomes proximal, lips elongated and spirally coiled ( Fig. 3A View Fig ). Gonoducts usually small, empty [somewhat elongated and filled with eggs or opaque white matter (sperm?) in specimens collected in August and early September (see Fig. 5 View Fig )]. Neurointestinal vessel issuing from ventral vessel (running along ventral nerve cord), forming a very narrow loop around interbasal muscle, subsequently dorsally bifurcated and terminating on each side of ring vessel ( Fig. 3A View Fig ). Rectal caecum present, but its connection with deteriorated ventral vessel unrecognizable. Paired anal vesicles, probably free (without mesenteries), but too damaged for detailed description.
Remarks. The present specimens were very similar to previous descriptions of L. brevirostris in having a very short proboscis, trunk terminal bulb, 7 longitudinal muscle bands, sickle-shaped ventral setae, interbasal muscle, two pairs of postsetal gonoducts, orientation of the neurointestinal vessel, and presence of a rectal caecum. Edmonds (1963) distinguished his newly-established L. bulbocaudatus from L. brevirostris by the presence (in the former) or absence (in the latter) of the terminal bulb, but later ( Edmonds 1987) found the bulb to be sometimes undetectable among specimens from the type locality of the former species ( Queensland, Australia), and concluded that the two species were conspecific ( L. brevirostris having priority). Listriolobus brevirostris is clearly distinguishable from the 5 congeners so far known (see Nishikawa 2004: table 3), including the only known Japanese congener L. sorbillans (Lampert, 1883) . The latter differs markedly from the Japanese specimens of L. brevirostris in the relative length of the proboscis (from 15% to more than 50% in the former vs. 4–28% in the latter), and in having usually 3 pairs of gonoducts ( vs. 2), 11–16 longitudinal muscle bands ( vs. 7), and the interbasal muscle absent ( vs. present). On the basis of previous descriptions and his examination of a paratype of Thalassema arkati Prashad, 1935 , Nishikawa (2004) suggested the possible conspecificity of L. brevirostris with T. arkati (type locality: Kolkata, India), given as Ochetostoma arkati by Biseswar (1988), and L. capensis (Jones and Stephen, 1955) ( South Africa), both with seven or eight longitudinal muscle bands and two post-setal pairs of gonoducts. Further, T. arkati was originally described as having structure similar to the terminal bulb in L. brevirostris (see Prashad 1935: pl. 1, fig. 1), although the above-referred paratype lacked it (Nishikawa, unpublished). Such a terminal structure has been reported also in South African specimens of O. formosulum (Lampert, 1883) by Biseswar (1988), and in the original descriptions of O. septemyotum DattaGupta, Menon, and Johnson, 1963 (from Quilon, SW coast of India) by DattaGupta et al. (1963), and Anelassorhychus dendrorhynchus ( Annandale and Kemp, 1915) (from Chilka Lake, E coast of India) by Annandale and Kemp (1915), though these generic affiliations do not seem to be firmly established to date. Morphological and molecular characteristics should be further addressed in future taxonomic revisions of L. brevirostris and its allies. Until then, the above-stated conspecificity remains unacceptable.
Among previous descriptions of L. brevirostris, DattaGupta and Menon (1966) provided the only account of mature specimens, collected in April, 1963 from Kerala, India. In the present study, mature specimens were collected in August and early September. Further collections of mature L. brevirostris from additional localities should cast more light on the biology of the species.
Geographical Distribution. See Fig. 2 View Fig : Osaka Bay, Osaka, Japan, 6.4–17.5 m deep, mud (present study); Bohai Bay ( Peng et al. 2013; Xiong et al. 2014; Wu et al. 2014), Kiaochow (=Jiaozhou) Bay, Shangtung ( Chen and Yeh 1958; Zhou et al. 2007), Lienyuan (= Jiangsu) Bay, Kiangsu ( Chen and Yeh 1958), Haimen Bay, Guangdong ( Li et al. 1997), Daya Bay, Guangdong ( Du et al. 2011; Zhang et al. 2017; Chen et al. 2019), and Baimajing, NW Hainan ( Li et al. 1994; Zhou et al. 2007), China; Hong Kong ( Kwok et al. 2008 based on Shin et al. 2004; Wang et al. 2017); Moreton Bay, Queensland, Australia, 8–10 fathoms ( Edmonds 1963); Yeppoon, Hervey Bay, and Bramble Bay, Queensland, Australia, mud ( Edmonds 1987); Cochin (= Kochi), Kerala, India ( DattaGupta and Menon 1966); Shivarajpur, Gujarat, India (Halder 2004); Kuwait Bay, Kuwait, Kuwait, 12 m (Al- Yamani et al. 2009).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Listriolobus brevirostris Chen and Yeh, 1958
| Nishikawa, Teruaki & Arase, Ikue 2019 |
Listriolobus aff. bulbocaudatus :
| Al-Yamani, F. Y. & Boltachova, N. & Revkov, N. & Makarov, M. & Grintsov, V. & Kolesnikova, E. & Murina, G. - V. 2009: 25 |
Listriolobus bulbocaudatus
| Stephen, A. C. & Edmonds, S. J. 1972: 423 |
| Edmonds, S. J. 1963: 245 |
Listriolobus brevirostris
| Al-Yamani, F. Y. & Skryabin, V. & Boltachova, N. & Revkov, N. & Makarov, M. & Grintsov, V. & Kolesnikova, E. 2012: 114 |
| Zhou, H. & Li, F. & Wang, W. 2007: 147 |
| Wang, W. & Zhou, Z. & Li, F. 1995: 31 |
| Li, F. & Wang, W. & Zhou, H. 1994: 206 |
| Edmonds, S. J. 1987: 131 |
| Stephen, A. C. & Edmonds, S. J. 1972: 424 |
| DattaGupta, A. K. & Menon, P. K. B. 1966: 193 |
| Chen, Y. & Yeh, C. 1958: 270 |