
Polyophthalmus zhadanae, Parapar & Al-Kandari & Candás & Moreira, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5052.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:6B3BF392-AB36-47BB-B306-79161F76F805 |
|
DOI |
https://doi.org/10.5281/zenodo.5579399 |
|
persistent identifier |
https://treatment.plazi.org/id/E8128798-2C58-F763-FF51-8B20FB2BFC0D |
|
treatment provided by |
Plazi |
|
scientific name |
Polyophthalmus zhadanae |
| status |
sp. nov. |
Polyophthalmus zhadanae View in CoL sp. nov.
Figures 1−11 View FIGURE 1 View FIGURE 2 View FIGURE3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 , 13 View FIGURE 13 ; Table 1 View TABLE 1
Material examined ( Table 1 View TABLE 1 ). Type material. Holotype (Museo Nacional de Ciencias Naturales; MNCN 16.01/19108 ) ; 208 paratypes (Museo Nacional de Ciencias Naturales; MNCN 16.01/19098 to 16.01/19107 and 16.01/19109 to 16.01/19121 ) ; 9 paratypes (Kuwait Institute for Scientific Research; no reference numbers) ; 33 paratypes (Seckenberg Museum; SMF30263 to SMF30267 ) ; 57 paratypes (Zoological Museum of Moscow State University—White Sea Branch; ZMMU-WS16458 to ZMMU-WS16461 ) ; 68 paratypes (Australian Museum; W.53384 to W.53387 ). Non-type material. 1,041 specimens ( KISR) . Additional material studied. Polyophthalmus pictus ( Dujardin, 1839) . 70 specimens ( MNCN 16.01/9402 , 16.01/13213 and MNCN 16.01/19122 to 16.01/19124 : Atlantic coast, Spain; MNCN 16.01/9375 , 16.01/9392 and 16.01/9398 : Mediterranean coast, Spain) .
Diagnosis. Parapodia biramous, chaetae emerging from body wall throughout; minute spherical dorsal and ventral parapodial cirri; inter-parapodial ciliated sensorial cilia present in all chaetigers and forming distinct lobes specially developed in anterior ones. Anal tube very short; posterior border with up to 6–7 pairs of digitate marginal papillae, of different length and often hidden inside anus.
......continued on the next page
......continued on the next page
......continued on the next page
......continued on the next page
......continued on the next page
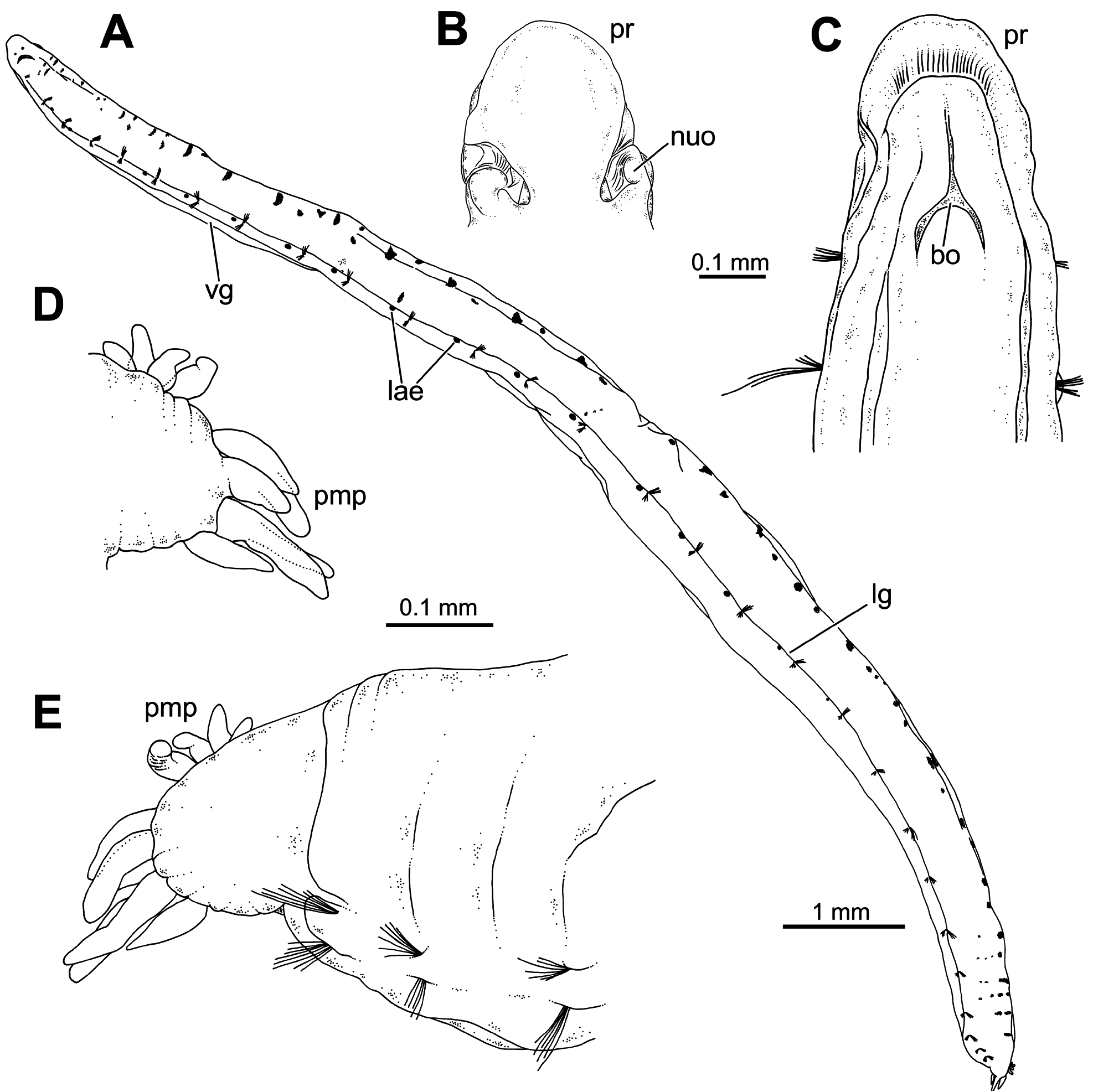
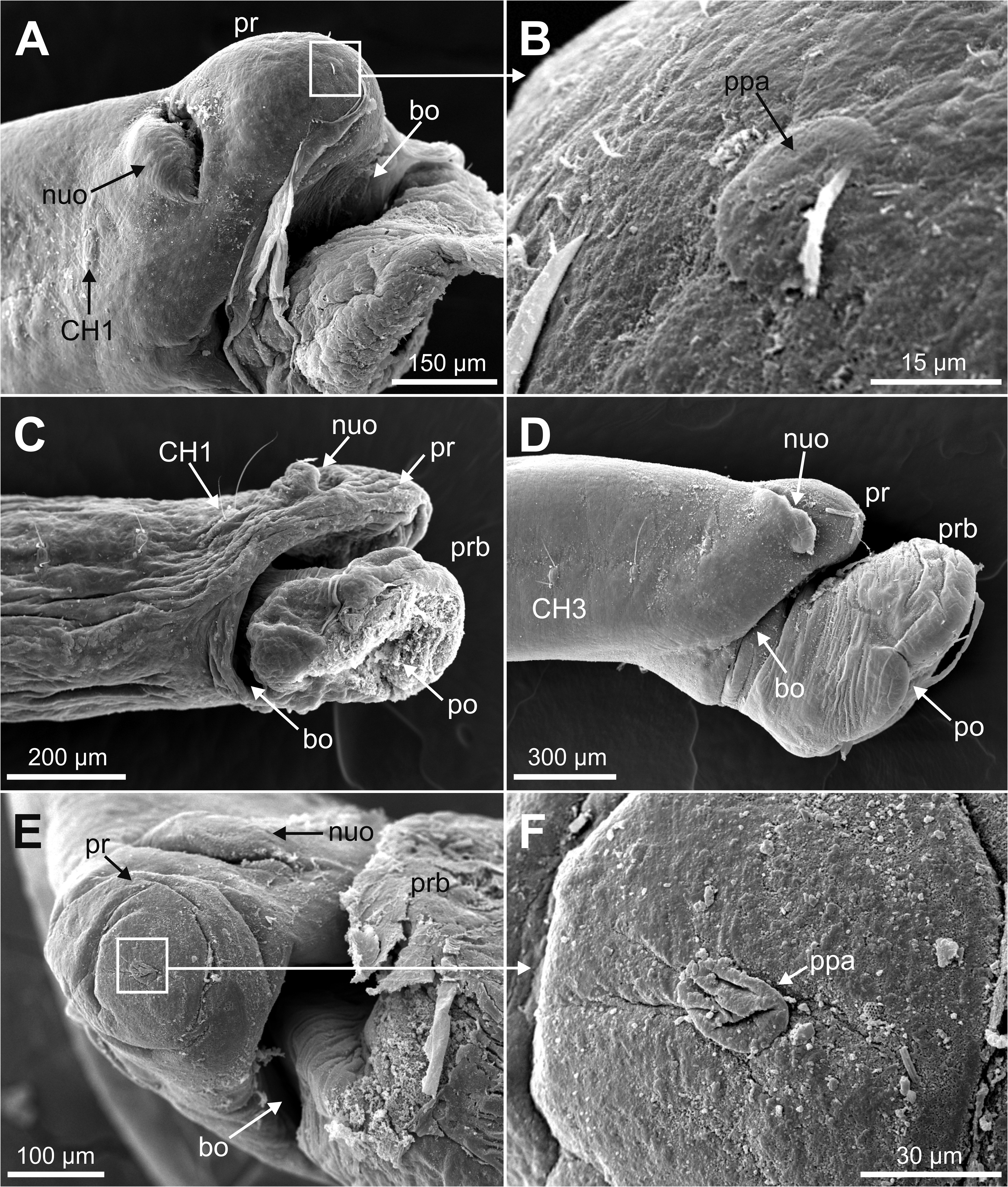
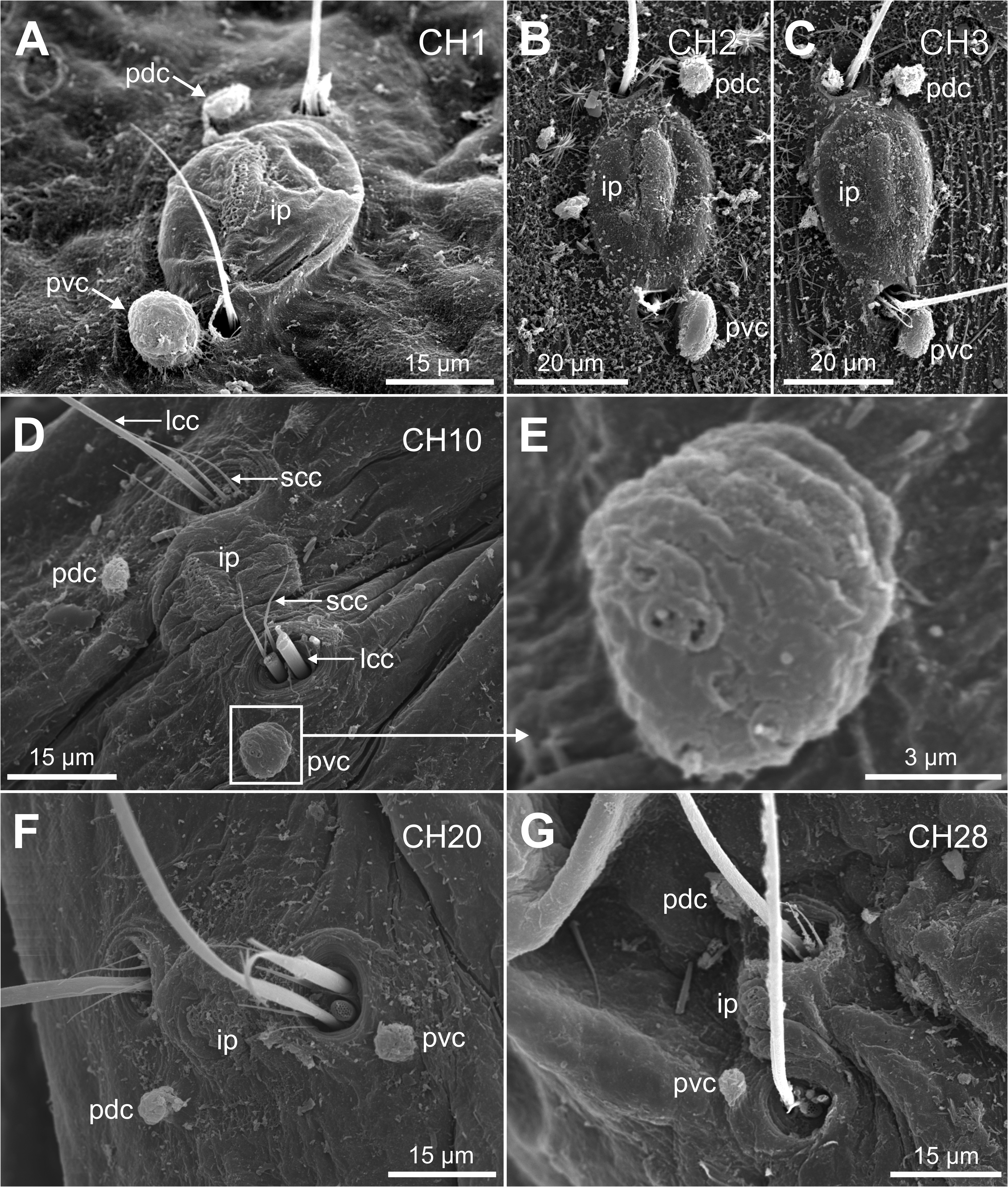
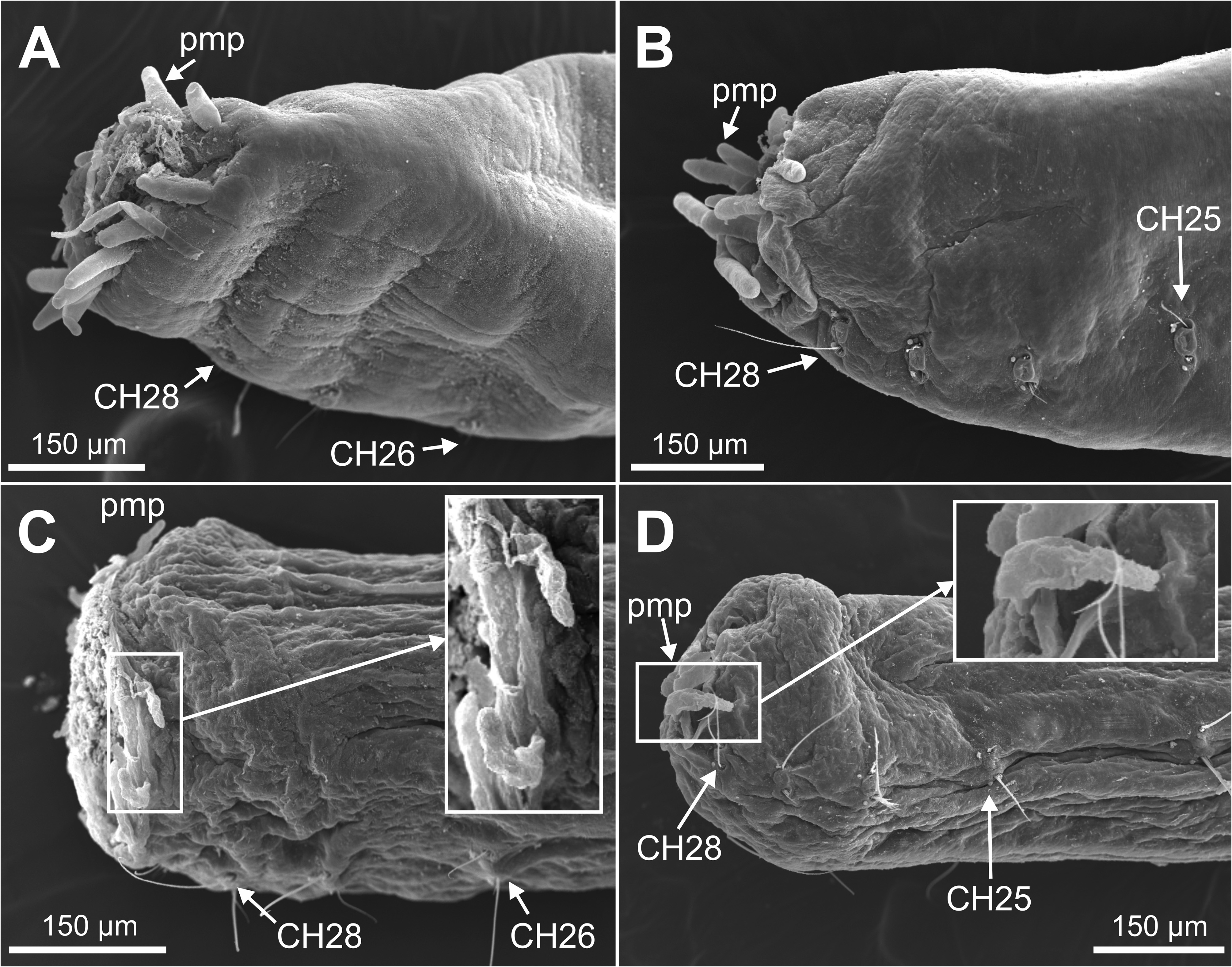
Description. Holotype complete specimen, 13.0 mm long and 1.0 mm wide for 28 chaetigers ( Fig. 2A View FIGURE 2 ); complete paratypes ranging from 10–12 mm in length and 0.5–1.0 mm in width, with up to 28 chaetigers. Body slender, tubular, tapering from last five chaetigers ( Figs 2A View FIGURE 2 , 3A, C View FIGURE3 ); lateral and ventral grooves from anterior to posterior body end ( Fig. 2A View FIGURE 2 ). Prostomium broadly rounded ( Fig. 2B–C View FIGURE 2 ), with a low papilla on distal end ( Figs 5A–B, E–F View FIGURE 5 , 8B View FIGURE 8 , 9A–B View FIGURE 9 , 10A View FIGURE 10 ). Nuchal organs large, as deep oval lateral depressions ( Figs 2B View FIGURE 2 , 3B View FIGURE3 , 5A, C–E View FIGURE 5 , 8C View FIGURE 8 , 10A–C View FIGURE 10 , 11C View FIGURE 11 ). Pharynx asymmetrical, dorsally lobate ( Figs 3D–E View FIGURE3 , 5C–D View FIGURE 5 , 8B View FIGURE 8 , 9A View FIGURE 9 ). Parapodia biramous, with minute sphaerical dorsal and ventral cirri; chaetae emerging from body wall ( Fig. 6A–G View FIGURE 6 ); interparapodial papilla with ciliated sensorial cilia between notopodia and neuropodial chaetae, forming a distinct lobe on first chaetigers ( Fig. 6A–C View FIGURE 6 ). Simple capillary chaetae arranged in two bundles: 1–2 long capillaries and 3–4 short capillaries per bundle through body ( Fig. 6A–D, F–G View FIGURE 6 ), short capillaries only discernable with SEM ( Fig. 6D View FIGURE 6 ). Chaetae of last posterior chaetigers slightly longer than anterior ones but not surpassing pygidium ( Figs 3C View FIGURE3 , 7B–D View FIGURE 7 ). Lateral eyespots anterior to parapodia on 12 chaetigers ( CH 7 to CH 18) ( Figs 2A View FIGURE 2 , 3E View FIGURE3 ); reddish brown in colour, rounded; first and last 1–2 pairs smaller than others ( Fig. 3E View FIGURE3 ). Anal tube very short, with dorsal and ventral notch; preserved specimens with posterior end showing different degrees of compression/retraction/tapering ( Figs 3C View FIGURE3 , 7A–D View FIGURE 7 ), appearing truncate in some specimens ( Fig. 7C–D View FIGURE 7 ). Posterior border provided with up to 6–7 pairs of digitate marginal papillae ( Figs 2D–E View FIGURE 2 , 7A–B View FIGURE 7 ) of different length, and often almost completely retracted into anal cavity ( Fig. 7C–D View FIGURE 7 ); basal papillae not observed, even in non-retracted anal tubes ( Fig. 7A View FIGURE 7 ).
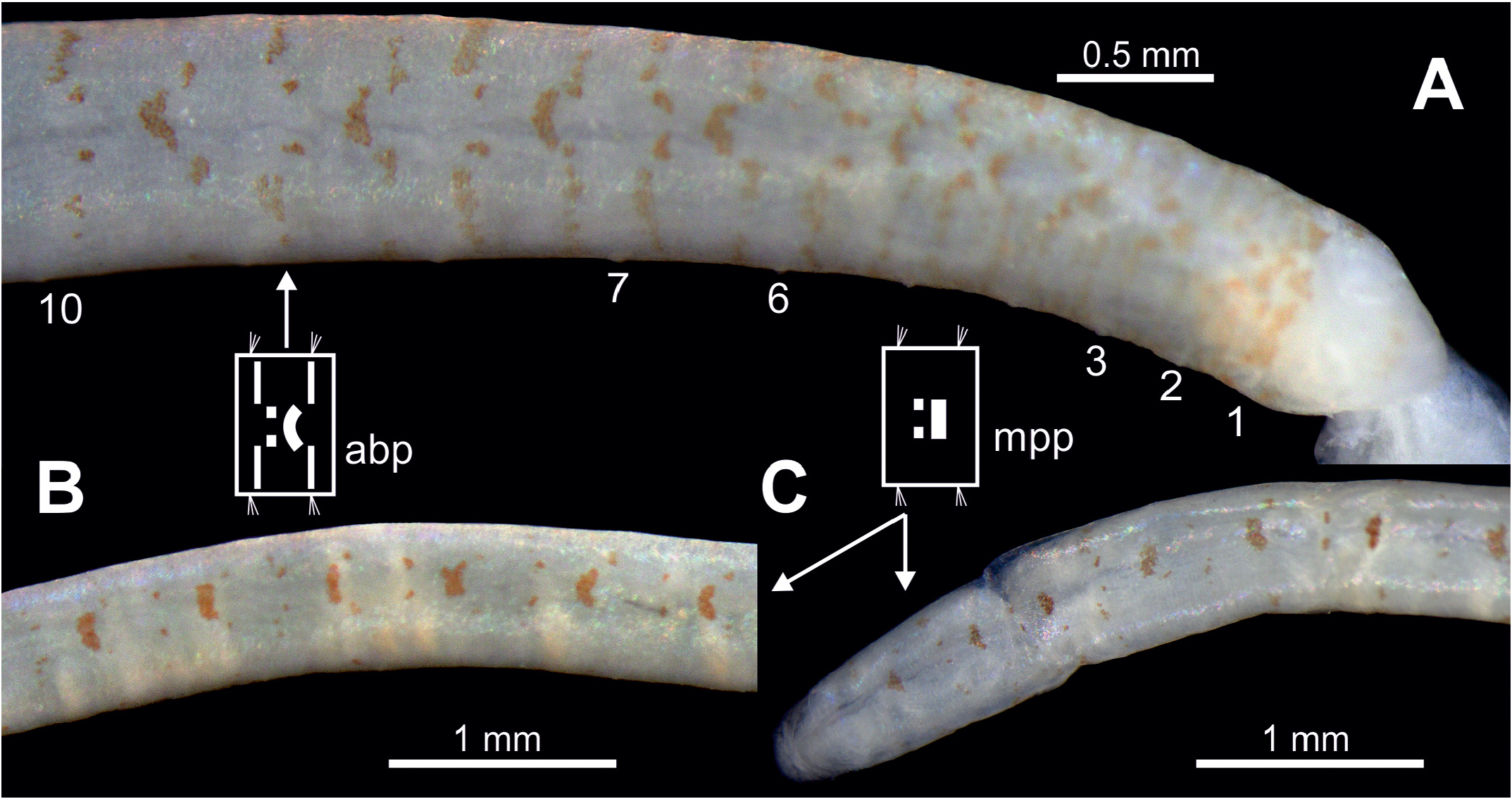
Holotype pale yellow in colour. Alive specimens with body pigmentation well-defined ( Fig. 3 View FIGURE3 ) but also even quite well preserved several years after fixation ( Fig. 4 View FIGURE 4 ) consisting of spots and stripes. Most specimens presenting a dorsal stripe (straight or slightly curved) and two brown spots behind per segment from midbody ( CH 10) to posterior end ( Figs 3C–D View FIGURE3 , 4B–C View FIGURE 4 ); holotype also showing two lateral stripes in CH 1. Other specimens presenting a similar pigmentation plus additional pigmentation from CH 1 to about CH 10: dorsal side of each chaetiger with one anterior half-moon shaped stripe with two spots behind and each lateral face with two stripes transversal to longitudinal body axis ( Figs 3B, D View FIGURE3 , 4A View FIGURE 4 ). Pigmentation absent ventrally in all specimens ( Fig. 3E View FIGURE3 ).
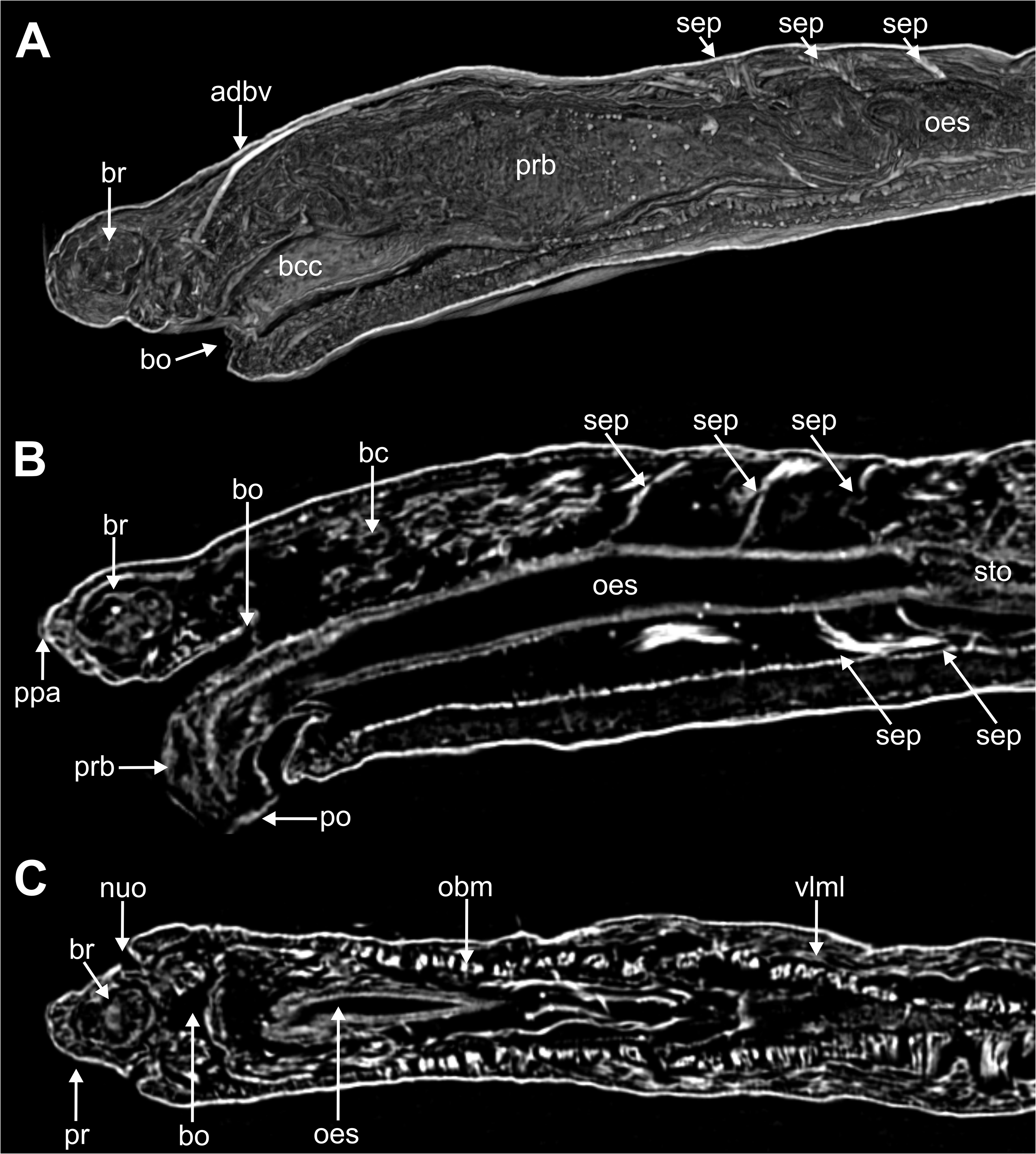
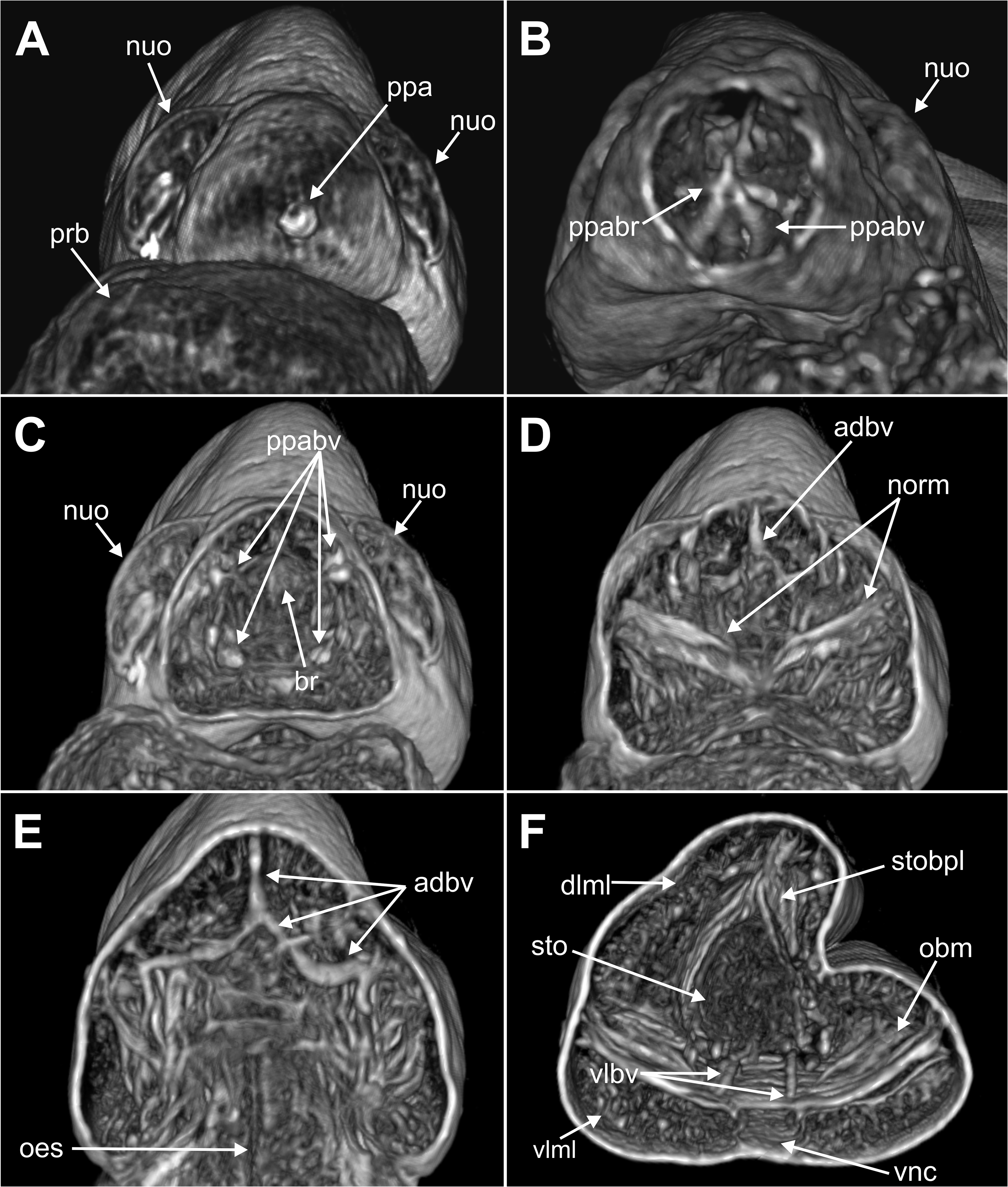
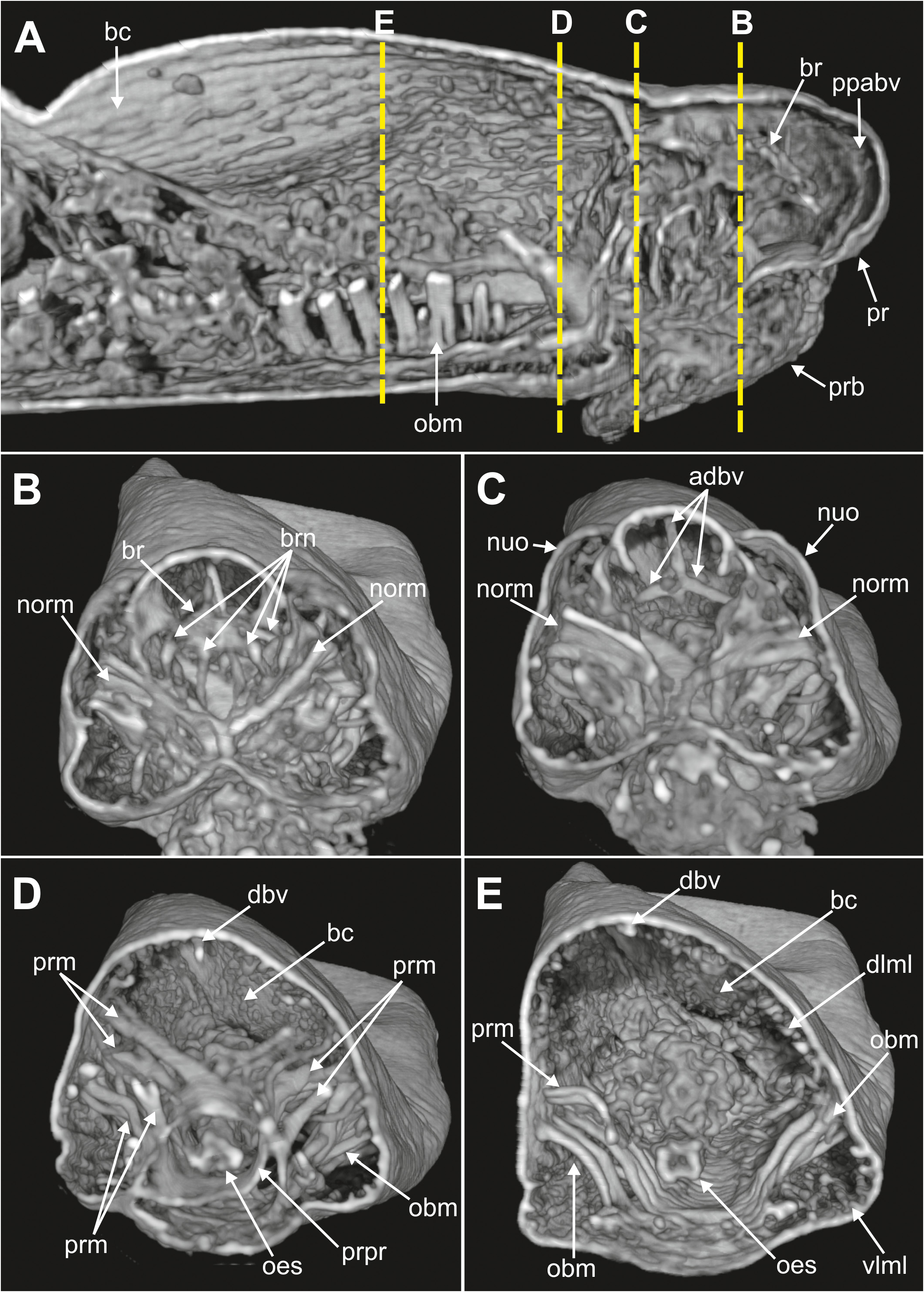
Internal anatomy. 2D and 3D Micro-CT images ( Figs 8–11 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 ) allowed the visualisation of the appearance and organization of (1) general body cavity and anterior septa, (2) anterior digestive tube, (3) musculature of body wall and associated to other structures, (4) circulatory and (5) nervous system. Other elements as nephridia and sensory organs could not be visualised.
(1) General body cavity is wide. It is particularly discernable in anterior body end when the proboscis is everted ( Figs 8B View FIGURE 8 , 9A–B View FIGURE 9 , 11A View FIGURE 11 ); the latter fills most of this space when retracted ( Fig. 8A View FIGURE 8 ). Septa are associated with proboscis protraction/retraction and first present at the level of oesophagus; they are visualised through sagittal 2D and 3D images whether the proboscis is retracted or not ( Fig. 8A–B View FIGURE 8 ).
(2) Foregut comprises a well-developed buccal cavity, proboscis, and oesophagus. The proboscis corresponds to the “asymmetrical dorsal lobed” type ( Figs 3E View FIGURE3 , 5C–D View FIGURE 5 , 8B View FIGURE 8 ). Oesophagus is connected to proboscis posterior end; its appearance seems convoluted when proboscis is retracted ( Fig. 8A View FIGURE 8 ) and straight when everted ( Fig. 8B View FIGURE 8 ); it is narrow in section and connected to septa associated to proboscis ( Fig. 8A–B View FIGURE 8 ); posterior end is connected to stomach and associated to a blood plexus ( Fig. 10E View FIGURE 10 ).
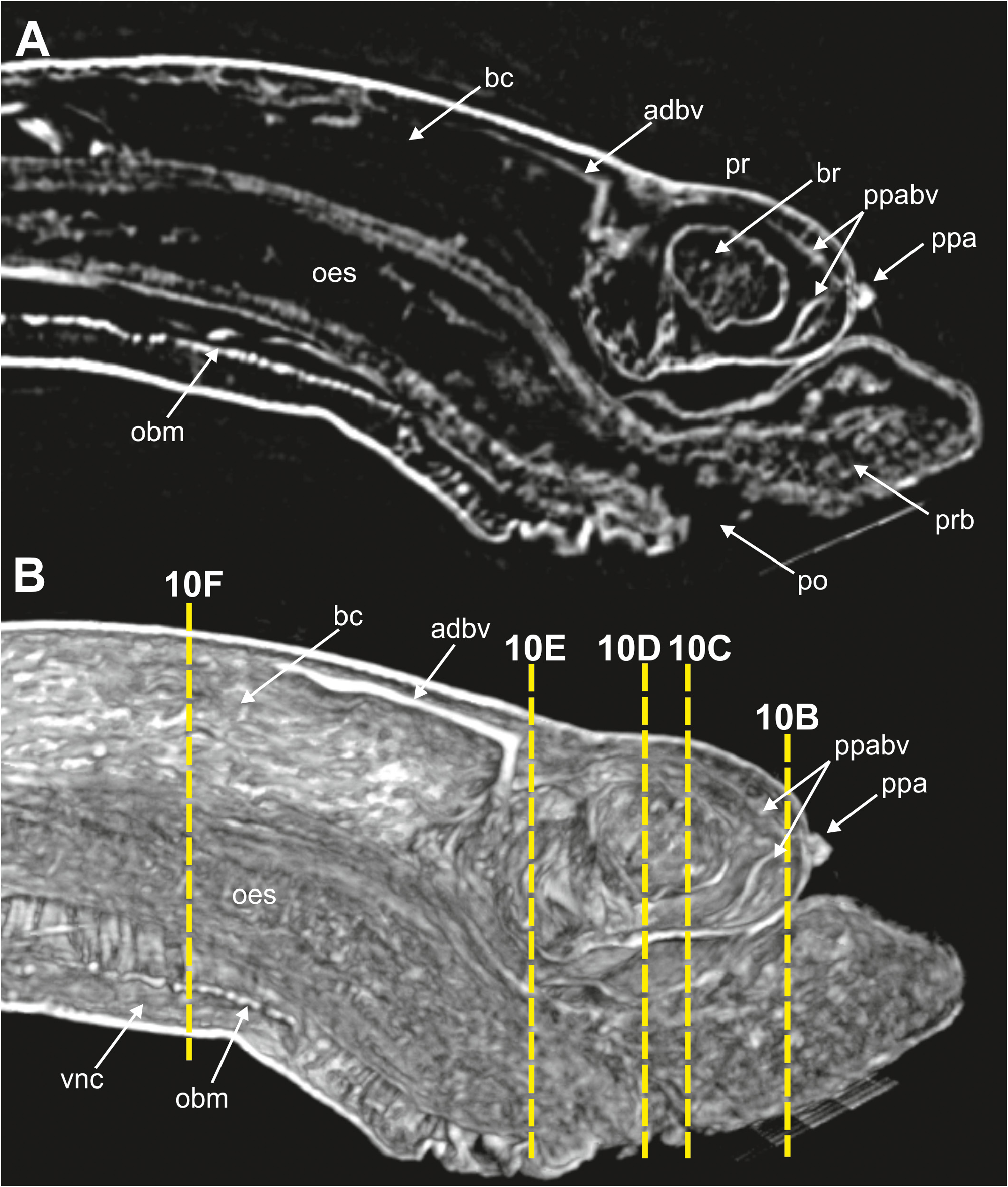
(3) Body wall muscles are organized in dorsal and ventral longitudinal bands ( Figs 8C View FIGURE 8 , 10F View FIGURE 10 ) and well-developed oblique bands, particularly in anterior body end ( Figs 8C View FIGURE 8 , 9 View FIGURE 9 , 10F View FIGURE 10 , 11A, D–E View FIGURE 11 ). Micro-CT images also clearly showed the retractor muscles of nuchal organs ( Figs 10D View FIGURE 10 , 11B–C View FIGURE 11 ) and those of proboscis ( Fig. 11D–E View FIGURE 11 ).
(4) Circulatory system was not visualised in full apart from the aforementioned blood plexus associated with stomach and two lateral longitudinal blood vessels located ventrally ( Fig. 10F View FIGURE 10 ); a longitudinal vessel seems extending forward near the prostomial papilla ( Figs 9A–B View FIGURE 9 , 10B View FIGURE 10 ) through a blood ring ( Fig. 10C View FIGURE 10 ) (see Discussion).
(5) Brain was clearly discernable ( Figs 8B–C View FIGURE 8 , 9A View FIGURE 9 , 10C View FIGURE 10 , 11A–B View FIGURE 11 ); ventral nerve cord corresponds to the space between the ventral longitudinal muscular bands ( Figs 9B View FIGURE 9 , 10F View FIGURE 10 ).
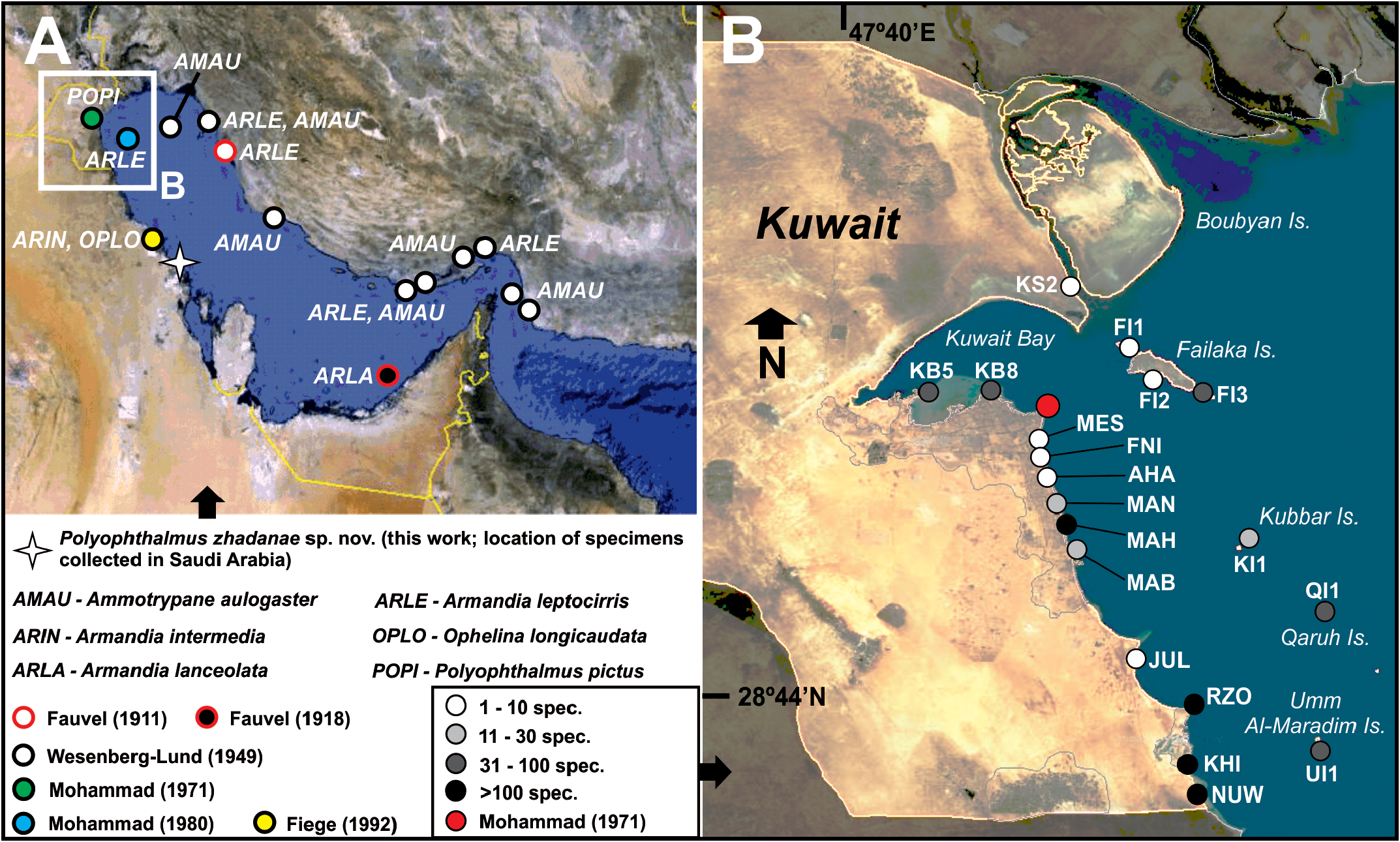
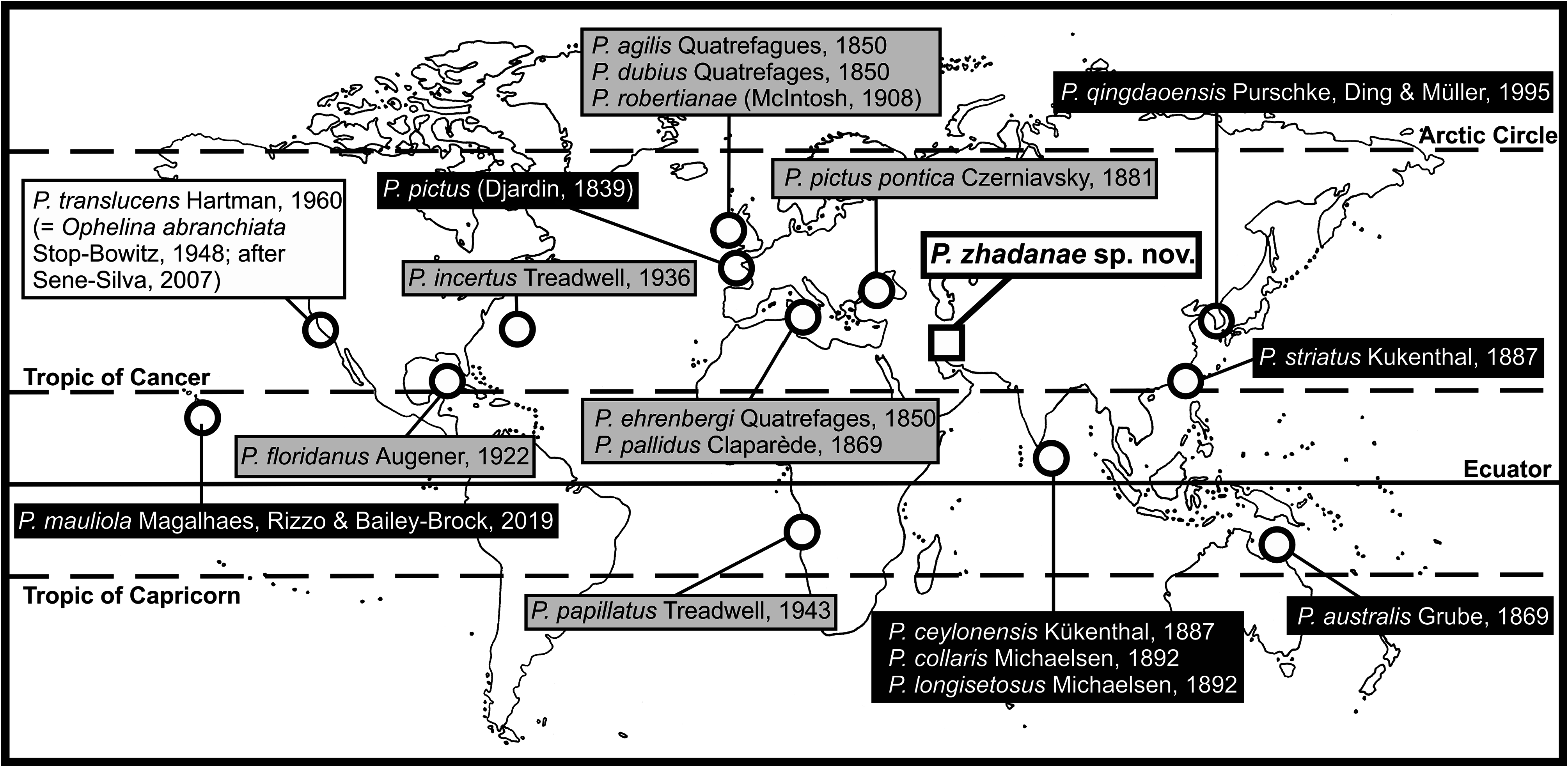
Type locality. Low intertidal in Ras Ajuza, East of Kuwait Bay , Kuwait; locality KB8, 29.39135ºN 47.99667ºE ( Fig. 1 View FIGURE 1 ; Table 1 View TABLE 1 ) GoogleMaps .
Distribution and bathymetry. Specimens have been collected in several intertidal localities of the coast of Kuwait and Saudi Arabia ( Fig. 1 View FIGURE 1 ), associated with different types of substrates ( Table 1 View TABLE 1 ).
Etymology. The new species is named after Dr. Anna Zhadan, White Sea Biological Station, Faculty of Biology, Lomonosov Moscow State University (Moscow, Russia), in recognition of her dedication to the study of the taxonomy of polychaetes and friendship.
Remarks. The main characters used to discriminate Polyophthalmus species are (see species key below): (1) pigmentation of prostomium/peristomium and body, (2) distribution and number of segmental eyespots, (3) presence/ absence of enlarged chaetae in posterior segments, and (4) shape and number of pygidial papillae. However, the usefulness of these characters is highly dependant on fixation techniques used, state of preservation of specimens and intraspecific variation. This is particularly true for body pigmentation (e.g., Fauvel 1927; Magalhães et al. 2019); examination of live specimens is therefore highly advisable ( Magalhães et al. 2019) but this is not always possible.
Magalhães et al. (2019) summarize the main morphological features of Polyophthalmus species described in the Indian and Pacific oceans, plus the Mediterranean P. pictus . Among them, P. zhadanae sp. nov. and P. mauliola Magalhães et al. (2019) are the only species supposedly provided with a ventral cirrus in all chaetigers; however, P. zhadanae sp. nov. bears an additional cirrus located dorsally in parapodia (absent in P. mauliola ) and interparapodial ciliated sensory lobe (“interramal papillae”) in all body parapodia while the latter are present only in the very posterior ones in P. mauliola .
Body pigmentation has been traditionally used as the main taxonomic discriminatory character for this genus ( Dujardin 1839), but apparently it shows a high intraspecific variability. This can be found in the type species of the genus, P. pictus , that shows quite different patterns of dorsal colouration (cfr. Dujardin 1839: Pl. 7, Figs 9–11 View FIGURE 9 View FIGURE 10 View FIGURE 11 vs. Fauvel 1927: Fig. 48o). We have observed such differences -as well as in relation to other highly taxonomic character: shape of the papillae of the anal tube- in specimens attributed to P. pictus from the Atlantic and Mediterranean coasts of the Iberian Peninsula ( Fig. 12A–E View FIGURE 12 ), even within the same population ( Fig. 12A View FIGURE 12 ). Anyway, the patterns of body pigmentation found in P. zhadanae sp. nov. makes this species close to P. ceylonensis Kükenthal, 1887 , that also shows transverse stripes at both sides in each of anterior five segments, and dorsal dark brown pigment patches on the dorsum of midbody to posterior segments (cfr. Figs 3C View FIGURE3 , 4 View FIGURE 4 -this work- vs Kükenthal 1887: Pl. 11, Fig. 10 View FIGURE 10 ). However, features of P. zhadanae sp. nov. pigmentation seems more complex across body. These species are also similar in body length (up to 11 mm in P. ceylonensis and 13 mm in P. zhadanae sp. nov.), number of chaetigers (27 in P. ceylonensis and 28 in P. zhadanae sp. nov.) and anal tube shape, that is short, open dorsally and ventrally (with 6 pairs of papillae in P. ceylonensis and 6–7 in P. zhadanae sp. nov.). Magalhães et al. (2019) report that the anal tube of P. ceylonensis is provided with two basal papillae and an unknown number of marginal papillae, and that the shape and length of the anal cone was “not described”. However, as explained above, Kükenthal (1887) described these characters; it is also likely that Magalhães et al. (2019) mentioned the presence of two basal papillae because of examination of the original figure as shown in ventral view (cfr. Kükenthal 1887: Pl. 11, Fig. 11 View FIGURE 11 ), that does not allow for adequate illustration of marginal papillae. Despite these similarities, P. ceylonensis and P. zhadanae sp. nov. differ in (1) the presence of a more complex pattern of pigmentation in P. zhadanae sp. nov., (2) the multiple annulations of body segments in P. ceylonensis , and (3) the lower number of segmental lateral eyes (9 pairs) in P. ceylonensis that are also of similar size, while P. zhadanae sp. nov. bears 12 pairs being the first and last pairs smaller than the others.
Other species with a similar number of segmental eyes are P. collaris Michaelsen, 1892 and P. australis Grube, 1869 (11 pairs) and P. qingdaoensis Purschke, Ding & Müller, 1995 and P. striatus Kükenthal, 1887 (12 pairs). However, body pigmentation in P. collaris is restricted to prostomium and peristomium, P. australis bears one pair of long basal pygidial papillae, P. qingdaoensis shows about 10 dark pigmented structures inside the brain including ocelli that are absent in P. zhadanae sp. nov., and P. striatus only bears four pairs of marginal pygidial papillae plus one basal pair also lacking dorsal and ventral parapodial cirri (see Table 1 View TABLE 1 in Magalhães et al. 2019).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |