
Eremidrilus pinedai, Rodriguez & Fend, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5159.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:132E04AE-187D-42AF-B0A7-68B4DCD7D0A9 |
|
DOI |
https://doi.org/10.5281/zenodo.6777140 |
|
persistent identifier |
https://treatment.plazi.org/id/72B42E3C-094F-41AB-BE7A-8B9D9D675AAF |
|
taxon LSID |
lsid:zoobank.org:act:72B42E3C-094F-41AB-BE7A-8B9D9D675AAF |
|
treatment provided by |
Plazi |
|
scientific name |
Eremidrilus pinedai |
| status |
sp. nov. |
3.3. Eremidrilus pinedai View in CoL n. sp.
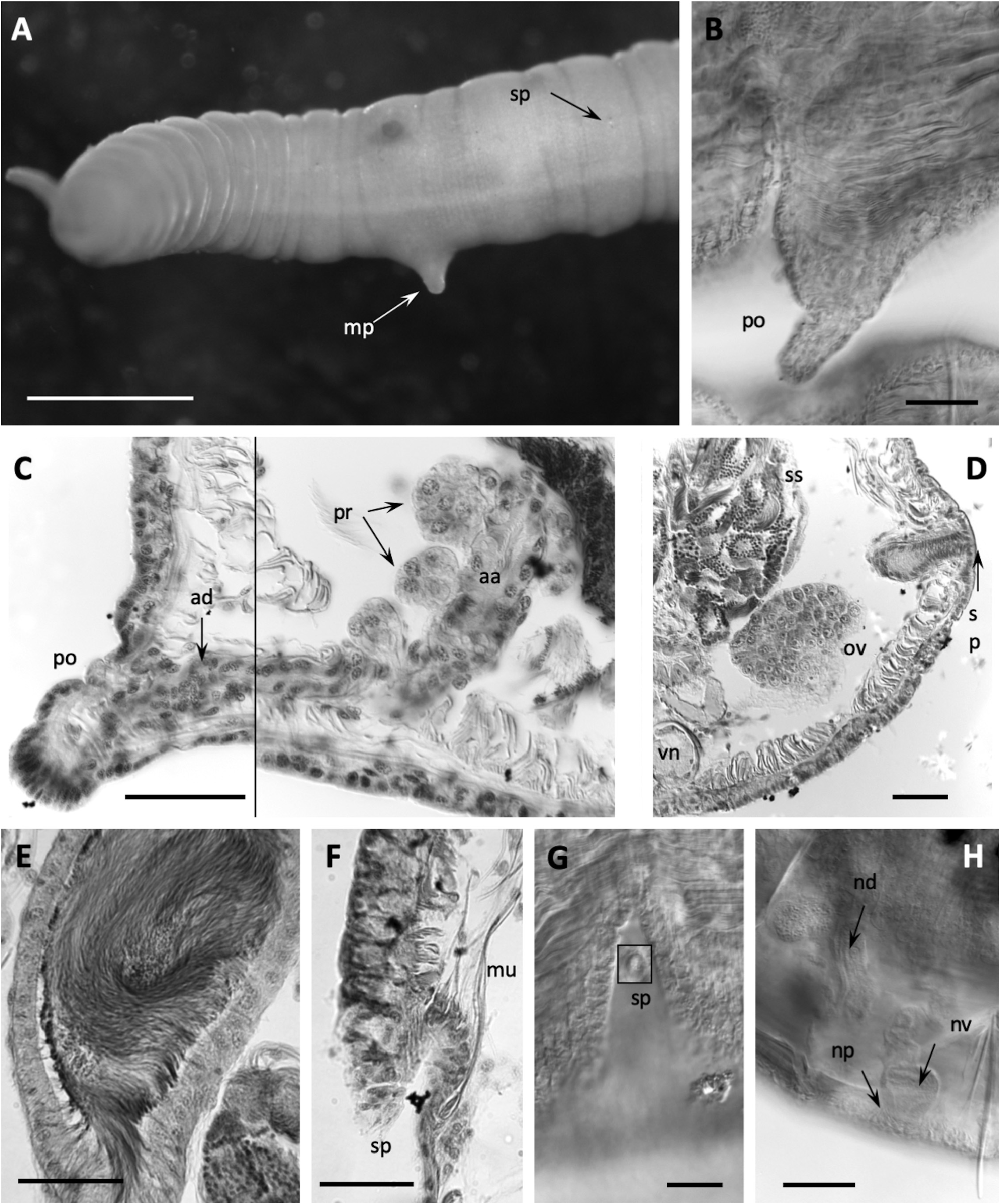
( Figs. 2B,E View FIGURE 2 ; 3F–I View FIGURE 3 ; 5 View FIGURE 5 )
Holotype. USNM 1593356 About USNM , whole worm, slide-mounted in Canada balsam.
Type locality. California, Del Norte Co., Smith River below forks, N 41.8729, W 124.1243, 40 m altitude (25 May 2002), S. V. Fend coll. GoogleMaps
Paratypes. USNM 1593357-1593360 About USNM , from the type locality, 25 May 2002, 2 whole-mounted, 1 dissected, 1 transversely sectioned. CASIZ 233662 , from the type locality, 25 May 2002, 1 whole-mounted; CASIZ 233663 from the type locality, 8 June 2003, 1 dissected on slide. MNCN 16.03 About MNCN /3149, from type locality, 25 May 2002, 1 whole-mounted, stained in borax carmine and mounted in Kleermount, and MNCN 16.03 About MNCN /3150 sagittally sectioned. S. V. Fend coll.
Etymology. Named after the naturalist Antonio Pineda, born in Guatemala, and head of the naturalist team on the Malaspina expedition (1789–1794) ( Madulid 1982), for his contributions to the knowledge of the Nearctic fauna.
Other material. From the type locality: 12 May 2001, 3 whole-mounted. 25 May 2002, 3 whole-mounted, 1 dissected, 3 sectioned (1 sagittal, 2 transverse). 8 Jun 2003, 1 whole-mounted, 1 dissected ( Table 1 View TABLE 1 ). S. V. Fend coll.
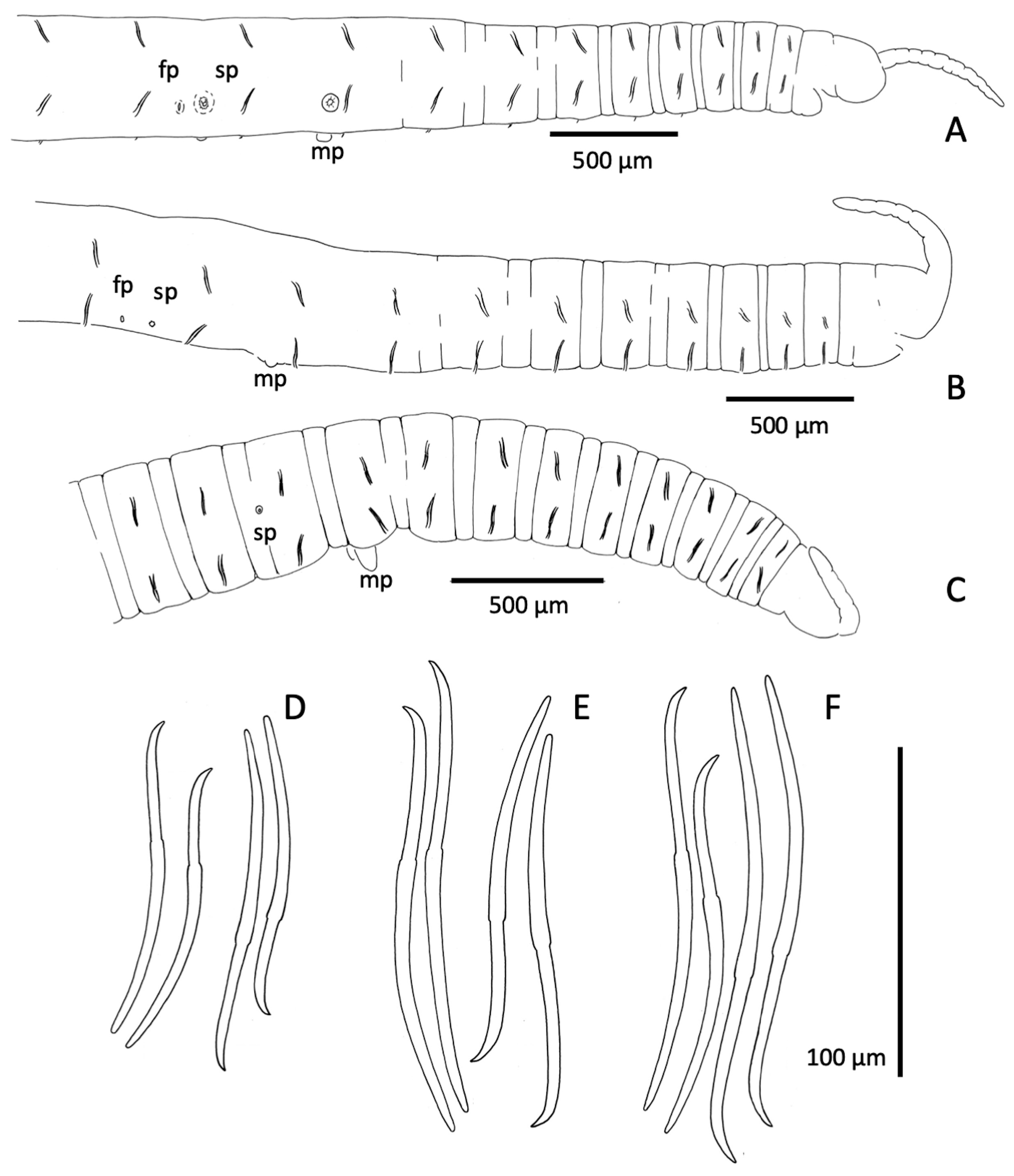
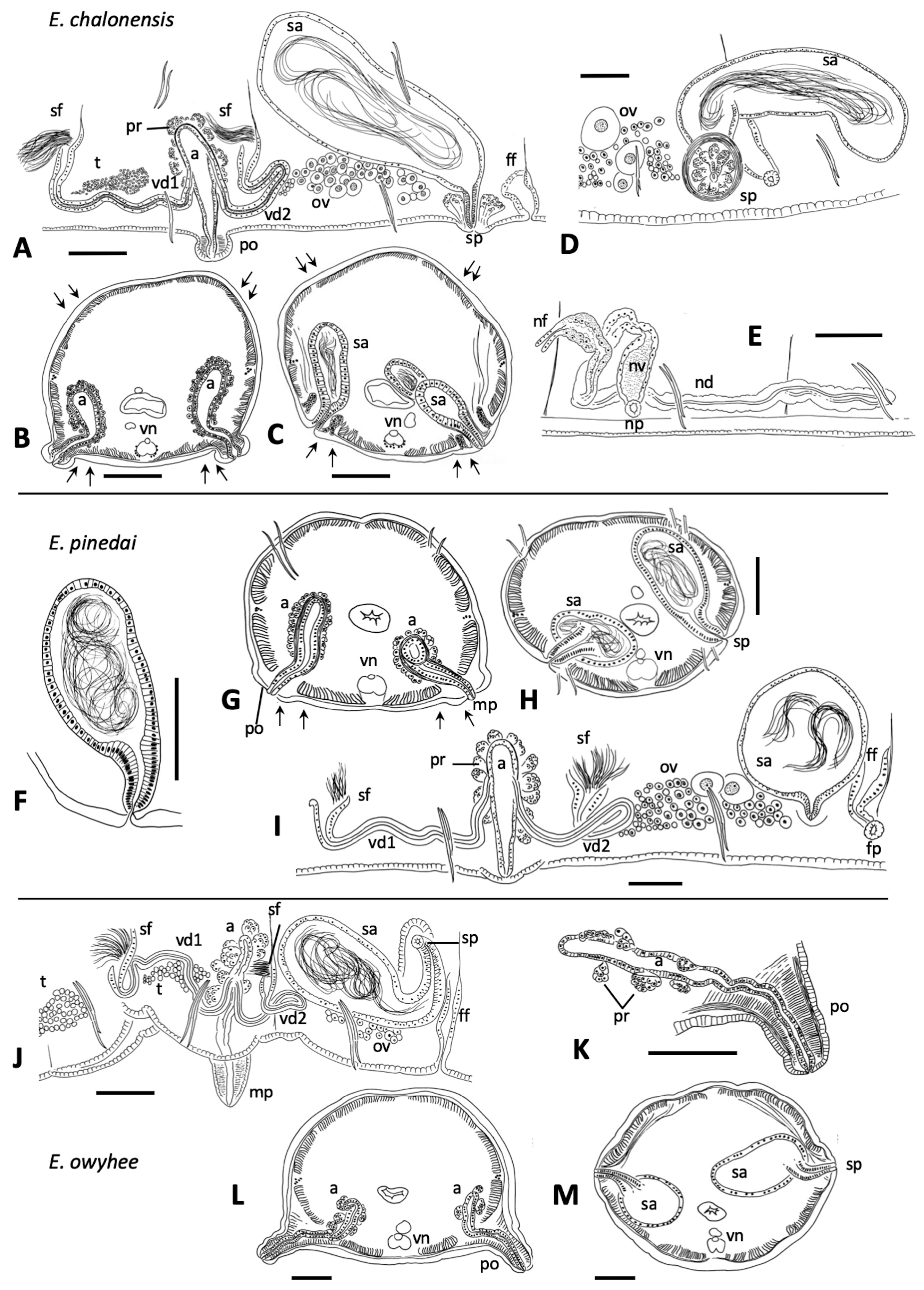
Description. Segments 60–98. Length of fixed worms 15–27 mm, body diameter in X 0.34–0.50 mm. Proboscis 418−675 µm long, about 70 µm diameter at middle ( Fig. 2B View FIGURE 2 ). Segmentation distinct in anterior segments; secondary annulus from IV to IX. All chaetae simple-pointed and sigmoid; in anterior body segments 113−155 µm long, shorter in II (ca. 65−75 µm), with nodulus slightly distal (about 0.4 chaeta length from the tip); in ventral bundles chaetae usually a little longer than dorsals of the same segment (median length of dorsals: 130 µm; ventrals: 134 µm) ( Fig. 2E View FIGURE 2 ). Chaetae in mid-body 108−167 µm long, only slightly shorter in posterior segments, and nodulus distal (0.3–0.4 from tip). Male pores on the line of ventral chaetae, on low and often indistinct porophores (18–32 µm high, 40–85 µm wide) located in the posterior part of segment X ( Figs. 2B View FIGURE 2 ; 3G, I View FIGURE 3 ; 5C, D View FIGURE 5 ). Spermathecal pores inconspicuous, opening in the ovarian segment, slightly lateral to ventral chaetae, and at about 2/3 of the distance from ventral chaetae bundle to posterior septum ( Figs. 2B View FIGURE 2 ; 3H, I View FIGURE 3 ; 5F View FIGURE 5 ). Female pores in 11/12, on line of ventral chaetae.
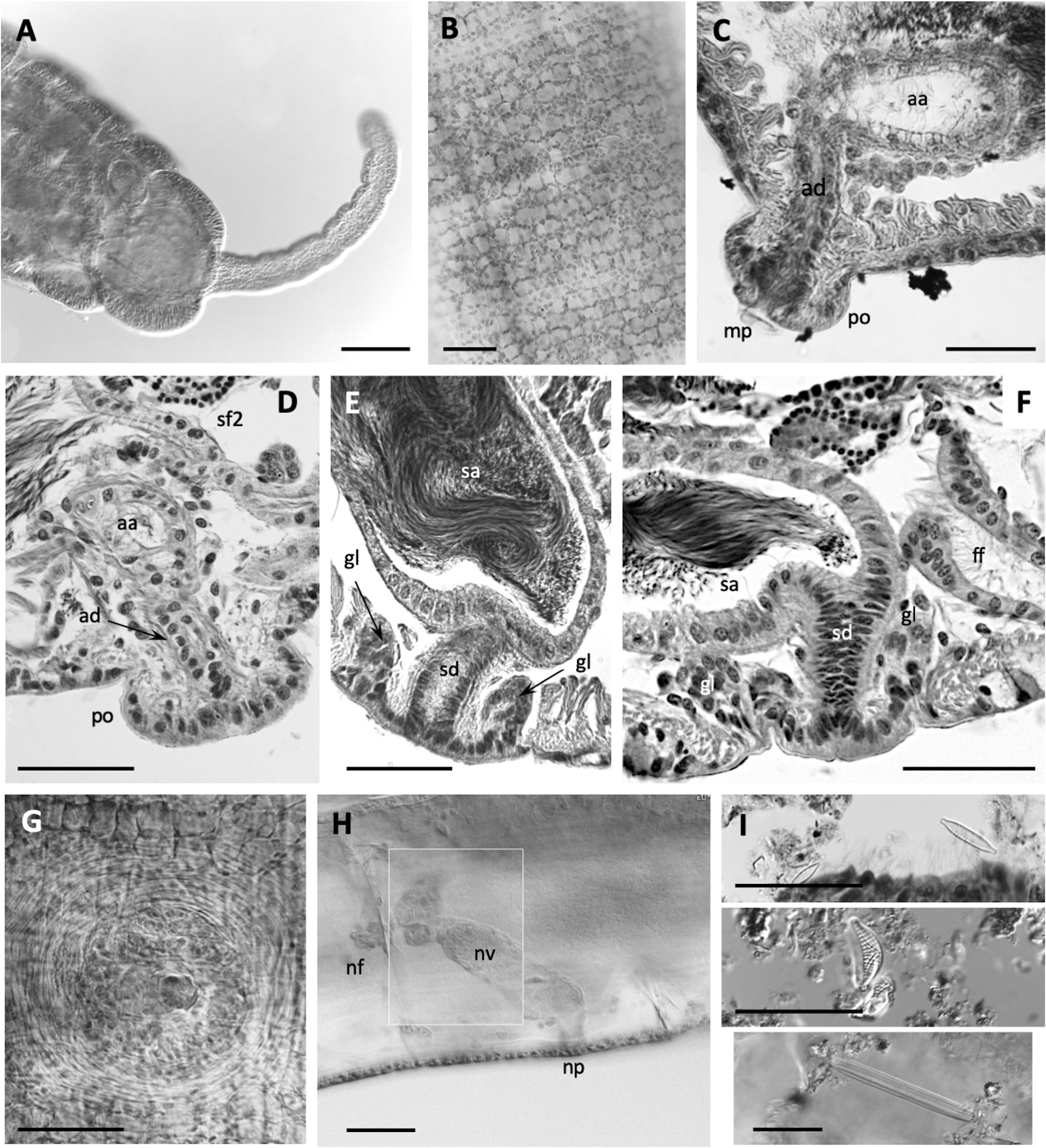
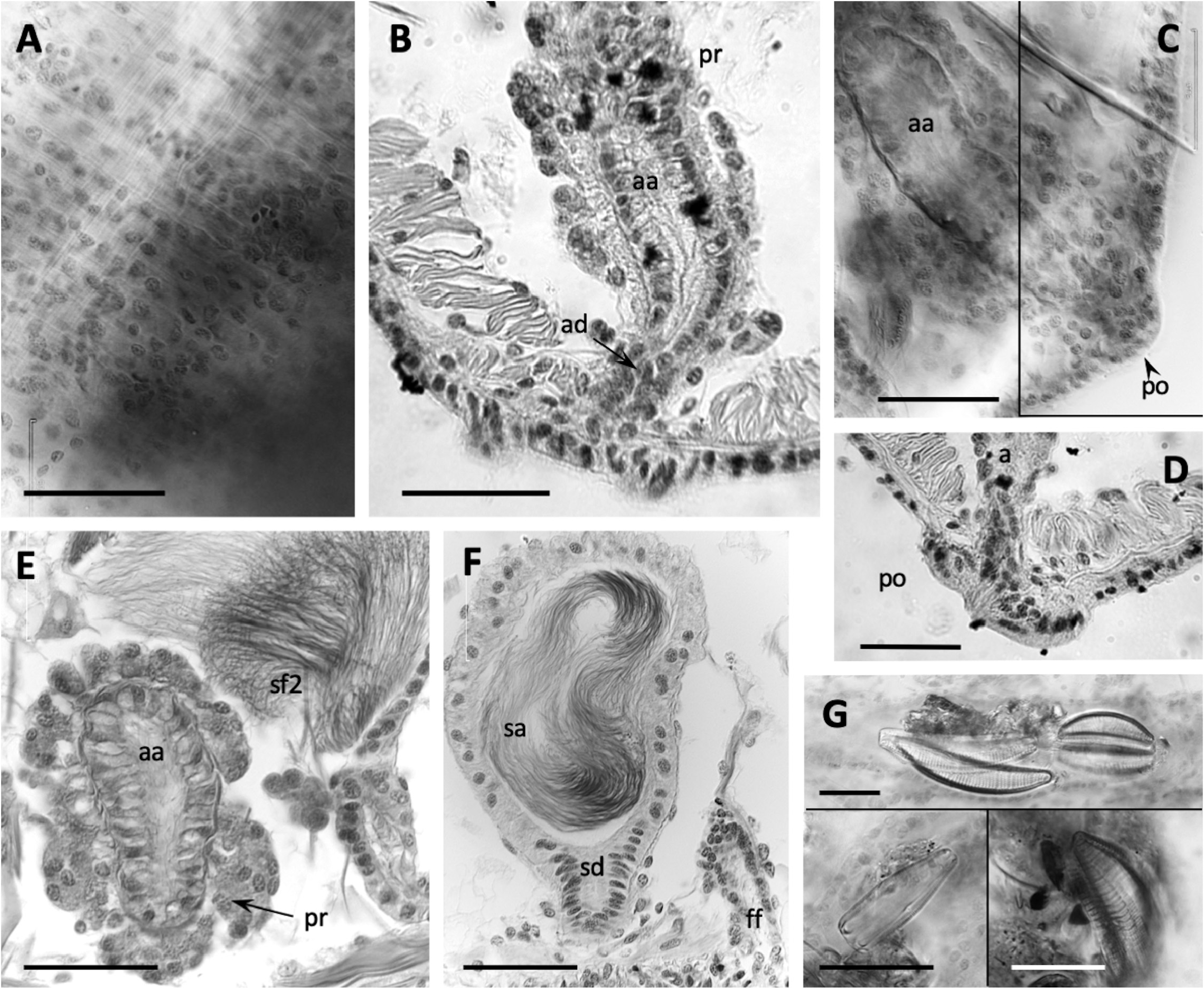
Epidermis in anterior segments 6–10 µm thick. Body diameter greatest in clitellar segments, but clitellum weakly developed in most mated specimens (8–10 µm thick); up to 16 µm thick in a few specimens, with small glandular cells aligned in transverse rows ( Fig. 5A View FIGURE 5 ). Longitudinal muscles to about 25 µm thick. Pharynx with a well-developed dorsal pad. Pharyngeal glands in (IV)V−VI(VII). Nephridia typical for the genus, composed of a short anteseptal funnel and tubular postseptal ducts that can extend through several segments dorsally and posteriorly; the ectal ducts about 10 µm diameter, without a terminal vesicle.
Sperm sacs usually extend anteriorly to VIII, posteriorly to XV; egg sacs to XVI. Vasa deferentia 14−24 µm wide and about 300−420 µm long, posterior about as long as anterior. Posterior vasa form a loop in XI, and both vasa join the atrial muscle layer near mid-atrium, entering the atrial lumen subapically. Atria entirely in X (148–244 µm long), club-shaped ( Figs. 3G, I View FIGURE 3 ; 5C View FIGURE 5 ), length about half the body diameter, muscle layer 2−4 µm thick, lining cells 6−12 µm high, and atrial ampulla densely covered by prostate glands (12–26 µm high), with densely-packed, often indistinct clusters ( Figs. 3G, I View FIGURE 3 ; 5B, E View FIGURE 5 ). Maximum atrium diameter 37–50 µm, narrowing towards male pore to about 20 µm just before the porophore. The ectal end of atrial duct may project slightly, forming a small papilla on the porophore ( Figs. 2B View FIGURE 2 ; 3G View FIGURE 3 ; 5C, D View FIGURE 5 ).
One pair of spermathecae opens in XI, and ampullae may extend into XII. Spermathecal ampullae ovate, 1.3 times longer than wide, about 5 times the length of the duct, and containing spermatozoids loosely clustered or forming a ring, but not oriented towards the epithelium ( Fig. 5F View FIGURE 5 ). Spermathecal ducts short and tapered (42–85 µm long, 32–48 µm maximum diameter, narrowing to about 20–30 at pore), length less than one fifth the body diameter, with muscle layer thin, and lining cells columnar ( Figs. 3H, I View FIGURE 3 ; 5F View FIGURE 5 ).
Remarks. As in other California species, E. pinedai n. sp. has only one pair of spermathecae, in the ovarian segment, but it is well distinguished by a combination of characters. As in E. chalonensis n. sp. (described above), the spermathecal duct is very short, i.e. the ratio of the spermathecal duct length to the body diameter in E. pinedai is 0.1–0.2; the spermathecal pore opens near the line of ventral chaetae, but it is not associated with conspicuous glands. The short spermathecal ducts and the ventral position of the spermathecal pores are reminiscent of the Eremidrilus species with two pairs of spermathecae (see Fend & Rodriguez 2020), but E. pinedai n. sp. is also distinguished by the small atria with short porophores. The atrium is short relative to the diameter of the body in X, as in E. humboldti and E. ritocsi . However, E. pinedai n. sp. has more weakly defined male porophores (cf. E. humboldti : Figs. 3A–E View FIGURE 3 ; 4A View FIGURE 4 , in Fend & Rodriguez 2020; E. ritocsi : Figs. 5C View FIGURE 5 6E View FIGURE 6 in Fend & Rodriguez 2003). In addition, E. humboldti has two spermathecal segments, and the spermathecal pores of E. ritocsi are midlateral. The male porophore of E. pinedai is smaller and usually less prominent than in most of the other Eremidrilus species having a single pair of spermathecae (cf. Figs. 9 and 12 in Fend & Rodriguez 2003) ( Table 2 View TABLE 2 ). The sperm organization within the spermathecae may also distinguish E. pinedai from E. ritocsi , in which sperm is arrayed in a characteristic spiral pattern (Fig. 11 in Fend & Rodriguez 2003). Dark detritus and a variety of diatoms were seen in the gut ( Fig. 5G View FIGURE 5 ).
The Smith River is an isolated, mid-sized (ca. 200,000 ha) drainage entering the Pacific Ocean in northwestern California. It is a large, alluvial stream without dams; discharge is highly variable, but flow is permanent ( National Water Information System, 2022). Much of the drainage is protected by the National Wild and Scenic Rivers Act (2022) and the river has large runs of several native salmonids ( Hogan & Zuber 2012). In addition to E. pinedai , the apparently endemic lumbriculid Kincaidiana smithi Fend & Rodriguez, 2017 ( Fend et al. 2017) was described from a nearby site. E. pinedai n. sp. was collected by digging into gravel/cobble sediments at the edge of a large gravel bar.
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |