
Echinoderes dubiosus, Sørensen & Rohal & Thistle, 2018
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.456 |
|
publication LSID |
lsid:zoobank.org:pub:DE1B1DEE-9871-4803-9F67-025F2B439560 |
|
DOI |
https://doi.org/10.5281/zenodo.3818846 |
|
persistent identifier |
https://treatment.plazi.org/id/312B7A46-B12C-49DE-B11B-D36EE2FCA488 |
|
taxon LSID |
lsid:zoobank.org:act:312B7A46-B12C-49DE-B11B-D36EE2FCA488 |
|
treatment provided by |
Valdenar |
|
scientific name |
Echinoderes dubiosus |
| status |
sp. nov. |
Echinoderes dubiosus sp. nov.
urn:lsid:zoobank.org:act:312B7A46-B12C-49DE-B11B-D36EE2FCA488
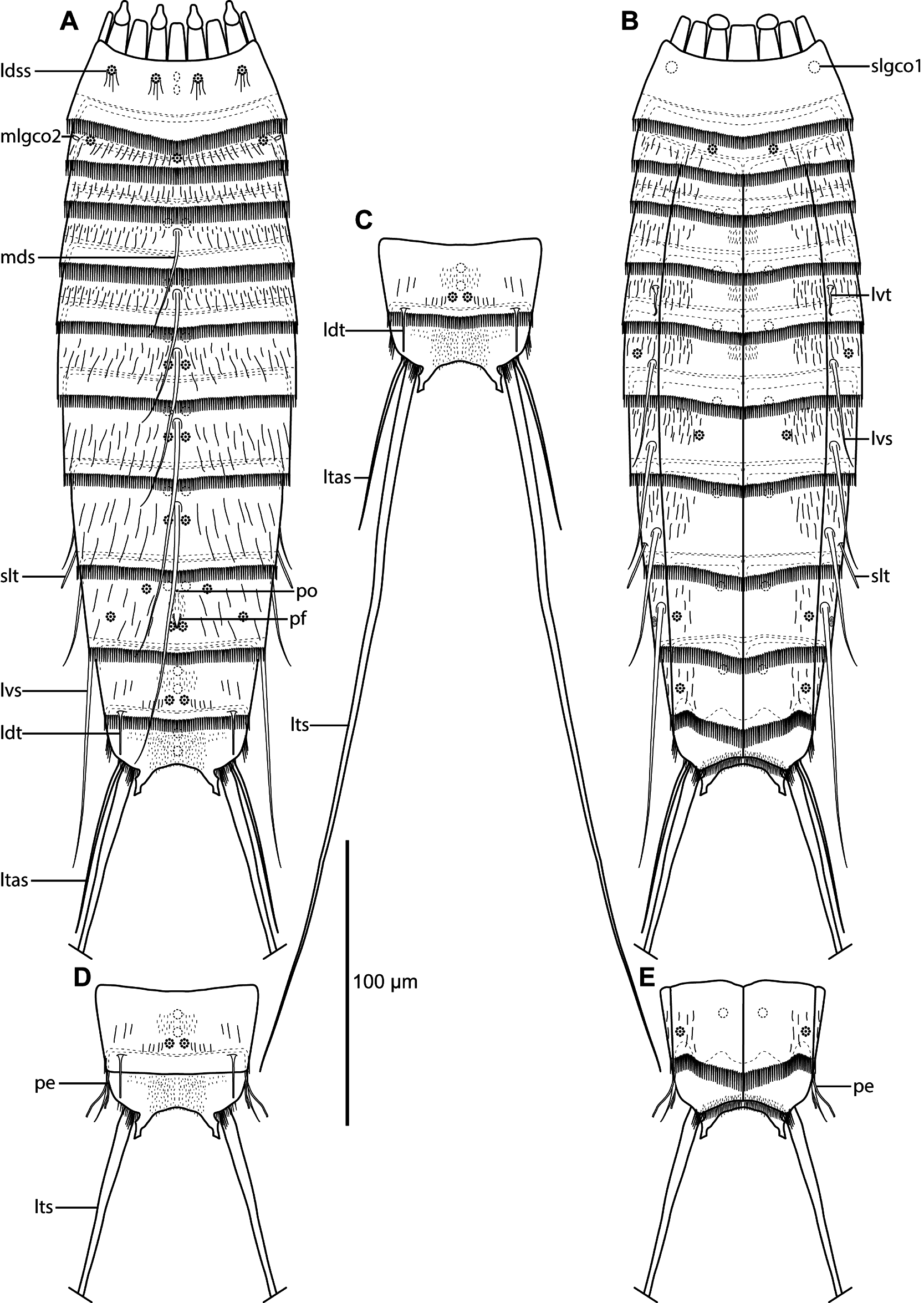
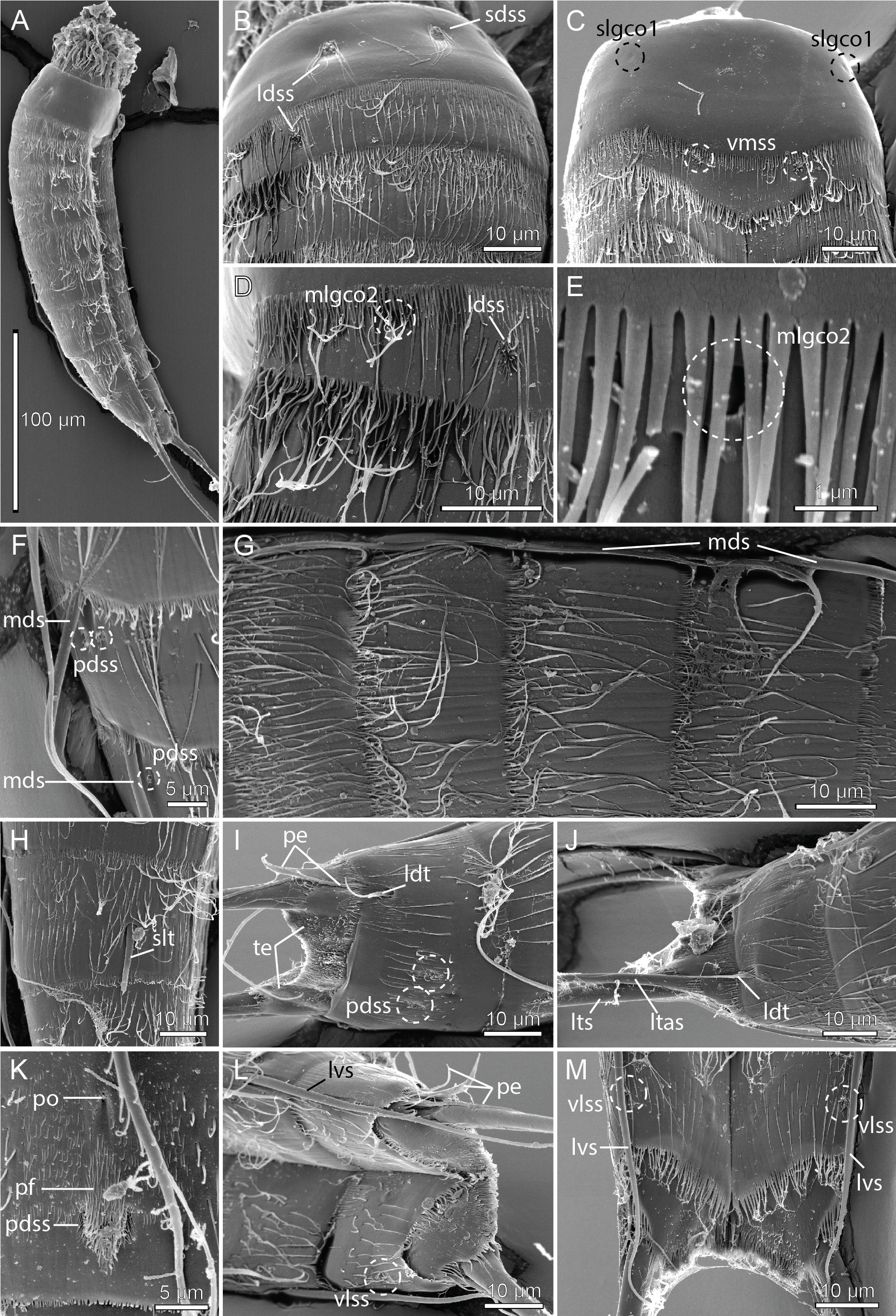
Figs 4 View Fig –6 View Fig View Fig View Fig , Tables 4–6 View Table 4
Diagnosis
Echinoderes with middorsal spines on segments 4 to 8 and spines in lateroventral positions on segments 6 to 9, with spines on segment 9 always extending beyond the terminal segment. Tubes present in lateroventral positions on segment 5, sublateral positions on segment 8 and laterodorsal positions on segment 10; tubes on segment 10 do not show sexual dimorphism. Minute glandular cell outlets type 2 present in midlateral positions on segment 2. Segment 9 with middorsal cuticular structures, forming an anterior pore, and a posterior papillary flap, flanked by two sensory spots. Males with three pairs of penile spines; females with lateral terminal accessory spines.
Etymology
This species name dubiosus is derived from the Latin ‘dubium’, because of its very close resemblance with Echinoderes bathyalis , and thus slightly questionable status as a distinct species.
Material examined
Holotype
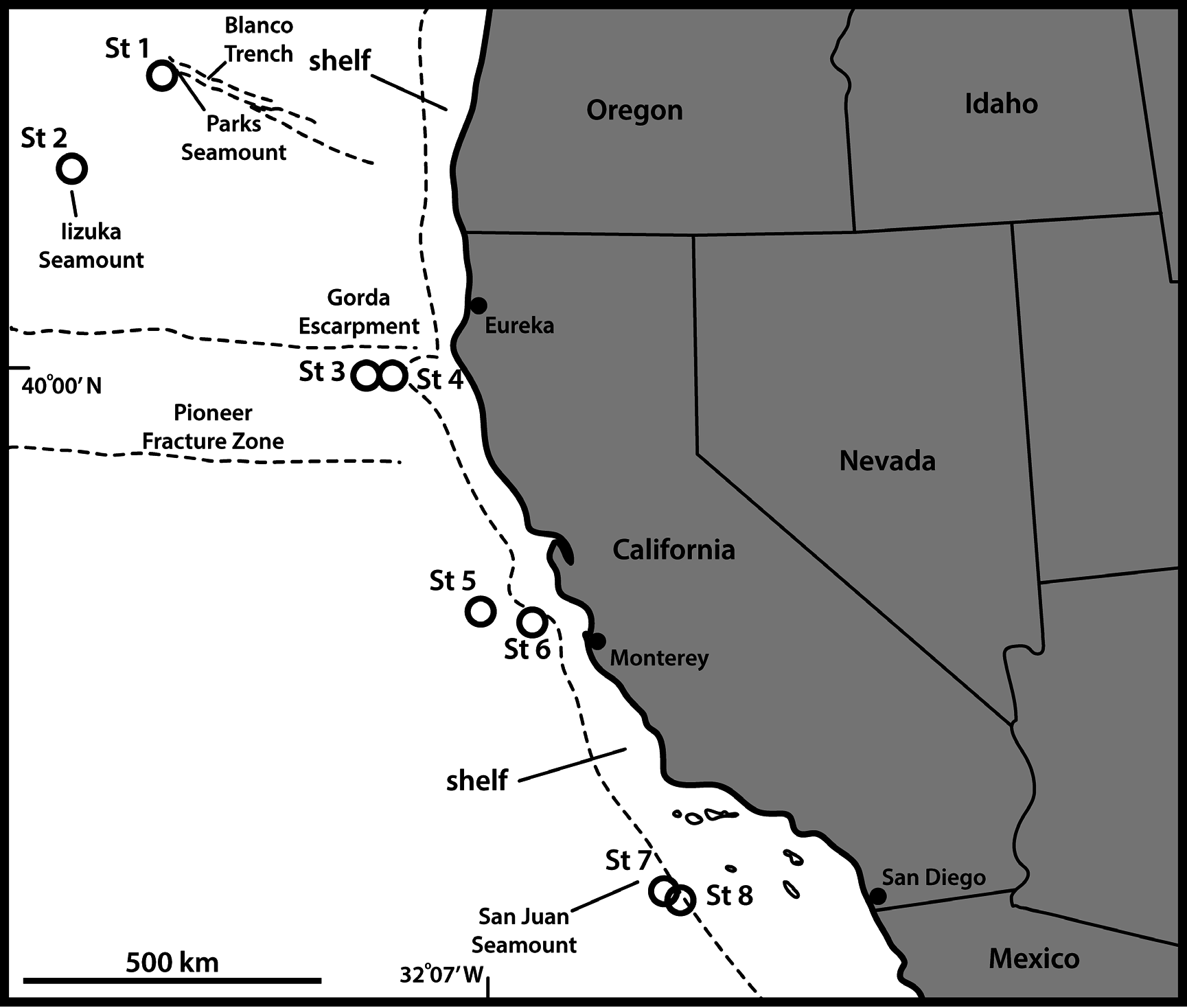
UNITED STATES OF AMERICA: adult ♀, US West Coast , off northern California, 39°59′58″ N, 125°52′27″ W, St. 3, 3675 m deep, collected from mud, 20 Sep. 2008, mounted on glass slide in Fluoromount G (NHMD-213583). See Fig. 1 View Fig for localities and Table 1 View Table 1 for detailed station data.
GoogleMapsParatypes
UNITED STATES OF AMERICA: 1 ♀, 1 ♂, same collecting data as for holotype; 2 ♀♀, St. 1; 1 ♀, St. 2; 1 ♀, St. 4; 2 ♀♀, St. 5; 1 ♂, St. 6; 2 ♀♀, 1 ♂, St. 8 (NHMD-213584–213595). All paratypes are mounted in Fluoromount G, nine on glass slides and three on H-S slides.
Additional non-type material
UNITED STATES OF AMERICA: 1 ♀, 1 ♂, St. 6; 1 ♀, St. 7. All mounted for SEM and stored in the first author’s personal reference collection.
Description
Adults with head, neck and eleven trunk segments ( Figs 4 View Fig A–B, 5A, 6A). The trunk appears relatively slender, with very long lateral terminal spines, often exceeding the trunk length ( Figs 4 View Fig A–C, 5A). For a complete overview of measurements and dimensions, see Table 4 View Table 4 . Distribution of cuticular structures, i.e., sensory spots, glandular cell outlets, spines and tubes, is summarized in Table 5.
None of the specimens mounted for SEM allowed a detailed examination of the head morphology. The neck has 16 placids, measuring 13 µm in length. The midventral placid is broadest, measuring 10 µm in width at its base, whereas all others are narrower, measuring 8 µm in width at their bases. Four dorsal and two ventral trichoscalid plates are present, with the ventral pair being broader than the two dorsal pairs.
Segment 1 consists of a complete cuticular ring ( Figs 4 View Fig A–B, 5B–C). Sensory spots are located medially on the segment in subdorsal and laterodorsal positions ( Figs 4A View Fig , 5B View Fig , 6B View Fig ), and two glandular cell outlets type 1 in middorsal positions ( Fig. 4A View Fig ), and one pair in sublateral positions ( Figs 4B View Fig , 6C View Fig ). The segment is completely devoid of cuticular hairs, except for long hairs around the sensory spots ( Fig. 6 View Fig B–C). The posterior segment margin is nearly straight, terminating in a pectinate fringe with thin, but quite long fringe tips ( Fig. 6 View Fig B–C).
Segment 2 consists of a complete cuticular ring, with minute glandular cell outlets type 2 located in midlateral positions ( Figs 4A View Fig , 5B View Fig , 6 View Fig D–E). Pachycyclus of the anterior segment margin is of regular thickness and uninterrupted. Sensory spots are located in middorsal, laterodorsal ( Fig. 6D View Fig ) and ventromedial ( Fig. 6C View Fig ) positions. Secondary pectinate fringe not detected on this or any of the following segments. On this and the eight following segments, the cuticular hairs are bracteate. The fairly long hairs are arranged in only two to three rows across the tergal plate ( Fig. 6B, D View Fig ) and on the lateral halves of the sternal plates. The posterior segment margin is nearly straight and consists of a pectinate fringe, with fringe tips being longer and thicker than those on preceding segment, and very flexible ( Fig. 6 View Fig B–D).
Segment 3, and remaining segments, consisting of one tergal and two sternal plates ( Figs 4 View Fig A–B, 5A, C, E–G). Pachycyclus of the anterior segment margin is well-developed and interrupted at the tergosternal and midsternal junctions, but also middorsally. No conspicuous cuticular structures present. Cuticular hairs and posterior segment margin straight as on preceding segment.
Segment 4 with acicular spine in middorsal position ( Figs 4A View Fig , 5B View Fig , 6G View Fig ). Sensory spots not observed. Anterior part of segment with glandular cell outlets type 1 in paradorsal and ventromedial positions. Pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 5 with acicular spine in middorsal position and minute tubes in lateroventral positions ( Figs 4 View Fig A–B, 5B–C, 6G). Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 6 with acicular spines in middorsal and lateroventral positions ( Figs 4 View Fig A–B, 5B, 6F–G). Sensory spots present in paradorsal ( Fig. 6F View Fig ) and sublateral positions. Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 7 with acicular spines in middorsal and lateroventral positions ( Figs 4 View Fig A–B, 5D, 6F–G). Sensory spots present in paradorsal ( Fig. 6F View Fig ) and ventromedial positions. Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 8 with acicular spines in middorsal and lateroventral positions ( Figs 4 View Fig A–B, 5D–E). Middorsal spine long, sometimes reaching or even exceeding the posterior margin of segment 11 ( Fig. 4A View Fig ). Rather thick and rigid tubes, without differentiated bases or lateral wings, present in sublateral positions ( Figs 5 View Fig E–F, 6F). The exact position of the tubes varied between the examined specimens; they always stay within the sublateral area, but in some specimens, including the holotype and some paratypes, the tubes are very close to the midlateral line, whereas in others the tubes are almost in lateral accessory positions (compare Fig. 5E and 5F View Fig ). The variation is not related to gender or population. Sensory spots are present in paradorsal positions only. Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 9 with acicular spines in lateroventral positions; the spines are thicker than other spines in the lateroventral series, and also long, always extending beyond the terminal segment ( Figs 4B View Fig , 5E View Fig ). A couple of uncommon cuticular structures are present in middorsal position ( Figs 4A View Fig , 6K View Fig ). Most anterior, it has an unpaired pore, flanked by cuticular ridges forming an inverted wedge. Posterior to this pore, a papillary flap or tuft of papillae, flanked by a pair of paradorsal sensory spots, is present. The structures are very conspicuous in SEM ( Fig. 6K View Fig ), but hardly visible in LM ( Fig. 5D View Fig ). Additional sensory spots present in subdorsal, laterodorsal and ventrolateral positions ( Figs 4 View Fig A–B, 5D). Small rounded sieve plates are present in sublateral positions ( Figs 4B View Fig , 5F View Fig ). Glandular cell outlets type 1, pachycycli, pectinate fringe of posterior margin and cuticular hairs as on preceding segment.
Segment 10 with laterodorsal tubes near posterior segment margin; tubes are long and thin, with bases and lateral wings in both sexes ( Figs 4A, D View Fig , 5H View Fig , 6 View Fig I–J). Sensory spots present in paradorsal and ventrolateral positions ( Figs 4 View Fig A–B, 6I, L–M). Glandular cell outlets type 1 present as two middorsal ones and one pair in ventromedial positions ( Fig. 5 View Fig E–F, H). Cuticular hairs scarcer than on preceding segments. The posterior segment margin of the tergal plate is straight; in females it terminates in a pectinate fringe with shorter fringe tips, whereas the fringe is hardly seen in males ( Fig. 6I View Fig ). The margins of the sternal ones are concave, extending posteriorly near the midventral junction ( Fig. 6M View Fig ); pectinate fringe with long fringe tips present in both sexes. Pachycycli as on preceding segment.
Segment 11 with lateral terminal spines, nearly equal in length to or longer than trunk length ( Figs 4C View Fig , 5A View Fig ). Males with three pairs of penile spines; dorsal and ventral penile spines are thin, flexible tubes, whereas the median ones are thicker and conical, yet flexible ( Figs 4 View Fig D–E, 5H, 6I, L); females with lateral terminal accessory spines ( Figs 4 View Fig A–B, 5G, 6J). No sensory spots observed. Glandular cell outlets type 1 present as two middorsal ones ( Fig. 5H View Fig ). The segment is completely devoid of cuticular hairs, but densely covered with short hair-like extensions, in middorsal to laterodorsal areas, and along the ventral margin ( Fig. 6I View Fig , L–M). Tergal extensions are short and pointed, with an extra tooth at the inferior margin ( Figs 4 View Fig , 5G View Fig , 6I View Fig ). Sternal extensions not present. Posterior margin of sternal plates with slightly longer fringe tips near the insertion of the lateral terminal spines.
Remarks
With middorsal spines on segments 4 to 8 and lateroventral tubes/spines on segments 5 to 9, E. dubiosus sp. nov. has the most common spine formula among species of Echinoderes . But still, the species is easily distinguished from most of its congeners by its thick and very long lateroventral spines on segment 9, combined with the presence of only midlateral glandular cell outlets type 2 on segment 2 and sublateral tubes on segment 8. Even though the spines on segment 9 usually are the longest in the lateroventral series of Echinoderes , they rarely reach the posterior margin of segment 10. Lateroventral spines of segment 9 extending beyond the posterior margin of the terminal segment is only known from E. dubiosus sp. nov., E. bathyalis and females of Echinoderes levanderi Karling, 1954 ( Yamasaki et al. 2018a; Sørensen 2018).
Moreover, the presence of glandular cell outlets type 2 in midlateral positions only on segment 2 is nearly unique among its congeners, and shared only with E. bathyalis . In general, whereas laterodorsal or sublateral glandular cell outlets type 2 on segment 2 are fairly common among species of Echinoderes , only two other species, E. aquilonius Higgins & Kristensen, 1988 and E. pennaki Higgins, 1960 , have glandular cell outlets type 2 in midlateral positions of this segment. However, these two species have 2 and 3 pairs of glandular cell outlets type 2 on segment 2, respectively, and additional pairs on segments 4, 5 and 8 (for E. aquilonius see Grzelak and Sørensen 2018; for E. pennaki see Herranz et al. 2018).
Hence, E. dubiosus sp. nov. is very easily distinguished from most congeners, except for E. bathyalis with which it shows a very close resemblance. Echinoderes bathyalis is also described from deep waters (2721–2875 m deep), but in the East Atlantic, near Sedlo Seamount, off the Azores. The two species have identical spine patterns, and E. bathyalis is also characterized by having long lateroventral spines on segment 9, extending well beyond the terminal segment. However, the middorsal spine of segment 9 in E. bathyalis appears to be longer than the one in E. dubiosus sp. nov., and always extending well beyond the terminal segment. In E. dubiosus sp. nov. it extends to the posterior margin of segment 11 only. Both species have tubes on segment 5, but, whereas they are present in lateroventral positions in E. dubiosus sp. nov., the tubes are slightly displaced to lateral accessory positions in E. bathyalis ( Yamasaki et al. 2018a) . Furthermore, unlike E. bathyalis , E. dubiosus sp. nov. also has sublateral tubes on segment 8. We would treat this latter difference with some caution though, because even though Yamasaki et al. (2018a) confirm the lack of such a tube in E. abyssalis , they note that one specimen has indications of a glandular cell outlet type 2 in lateral accessory positions on segment 8. Since the attachment sites of tubes sometimes resemble glandular cell outlets type 2, and the actual tubes may be difficult to visualize in LM, we cannot reject the possibility that some specimens of E. bathyalis have tubes on segment 8 in nearly the same positions as E. dubiosus sp. nov. During the preparation of the present description we contacted Dr Yamasaki regarding the nature of this structure in E. bathyalis and he confirmed that it was only observed in one specimen (out of three), that it seemed to be a glandular cell outlet type 2, rather than a tube, and that the position was clearly lateral accessory rather than sublateral (H. Yamasaki, pers. comm.). Hence, even though the differences are subtle, we would still consider them as diagnostic. Echinoderes dubiosus sp. nov. furthermore has laterodorsal tubes on segment 10. Such tubes are missing in E. bathyalis . However, Yamasaki (pers. comm.) could not confirm the absence of such tubes in E. bathyalis with complete certainty, because they sometimes are very difficult to see with LM. The two species also seem to differ from each other in their distribution of sensory spots though. Echinoderes bathyalis has no laterodorsal sensory spots on segment 2, but quite distinct subdorsal sensory spots on segment 3 ( Yamasaki et al. 2018a, and subsequently confirmed by Yamasaki, pers. comm.). SEM images of E. dubiosus sp. nov. quite clearly show the presence of laterodorsal sensory spots on segment 2 ( Fig. 6D View Fig ) and confirm their absence in subdorsal positions on segment 3 ( Fig. 6B View Fig ). Finally, E. dubiosus sp. nov. has an unusual papillary flap in middorsal position on segment 9. This structure seems to be absent in E. bathyalis , but as stressed above, the structure is very difficult to visualize with LM. According to Yamasaki (pers. comm.) a similar middorsal structure could “possibly be present” in E. bathyalis also.
Hence, in summary, E. dubiosus sp. nov. and E. bathyalis can be distinguished by the proportionally longer middorsal spine of segment 9 in E. bathyalis , and in differences in sensory spot positions on the tergal plates of segments 2 and 3. Additional potential differences are the presence/absence of tubes on segments 8 and 10, and a middorsal papillary flap on segment 9. Despite the subtle differences, we
would consider the two species to be distinct, but the resemblance clearly indicates that they are very closely related in spite of the geographic distance.
One of the noteworthy traits in E. dubiosus sp. nov. is the unusual presence of middorsal structures on segment 9. Middorsal structures in Echinoderes usually include acicular spines, glandular cell outlets type 1 and sensory spots. Some species also have an extending protuberance, but this structure would always be protruding from the intersegmental joint between segments 10 and 11 (see, e.g., E. augustae Sørensen & Landers, 2014 , E bookhouti Higgins, 1964 , and E. charlotteae Sørensen, Herranz & Landers, 2016 ( Sørensen & Landers 2014; Sørensen et al. 2016a)). Segment 9 of E. dubiosus sp. nov., however, has an unpaired, middorsal pore, flanked by cuticular ridges anterior to a papillary flap or tuft of papillae, which again is flanked by a pair of paradorsal sensory spots. Such an arrangement has not been reported from any other kinorhynch. We do know, however, about three other cases of uncommon middorsal structures among species of Echinoderes , and, interestingly, they also appear on segment 9. The description of E. gizoensis Thormar & Sørensen, 2010 reports on segment 9 the presence of a middorsal: “…elongate fringed area of filiform cuticular extensions, with a small pore”, situated between a pair of paradorsal sensory spots ( Thormar & Sørensen 2010). This structure somehow resembles the anterior middorsal pore in E. dubiosus sp. nov., even though the new species lacks the filiform cuticular extensions. But then again, the structure in E. gizoensis might also simply be a single-pored sensory spot with very long papillae. The other unusual middorsal structure on segment 9 was reported from E. levanderi . In the species’ recent redescription, Sørensen (2018) mentioned that a “..cuticular depression or scar, lined with short hairs is present in middorsal position, in between the paradorsal sensory spots”, and a nearly similar structure is found in E. yamasakii sp. nov. (see description below). Again, it is suggestive that this puzzling middorsal structure also appears on segment 9, and, as in the case of E. gizoensis and E. dubiosus sp. nov., is flanked by paradorsal sensory spots, but besides this, it is hard to argue for or against a homology between any of these structures. Sørensen (2018) was furthermore slightly reluctant about the nature of this structure because it was visible in SEM only, and only one SEM specimen was orientated in a way that allowed examination of the middorsal position of segment 9. The observations from the four species suggest though, that extra attention should be paid to similar structures in future descriptions.

| US |
University of Stellenbosch |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |