
Magelona boninensis, Taylor & Mortimer & Jimi, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5196.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:D11B689F-70DF-4B27-959D-63A520D125E2 |
|
DOI |
https://doi.org/10.5281/zenodo.7244459 |
|
persistent identifier |
https://treatment.plazi.org/id/3CED40AC-71DB-4DF0-AFA8-FB56B1862FF2 |
|
taxon LSID |
lsid:zoobank.org:act:3CED40AC-71DB-4DF0-AFA8-FB56B1862FF2 |
|
treatment provided by |
Plazi |
|
scientific name |
Magelona boninensis |
| status |
sp. nov. |
Magelona boninensis View in CoL sp. nov.
[Japanese name: Bonin-morote-gokai]
Figs 14–18 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 View FIGURE 18
Type locality: Bonin Islands, Japan
ZooBank LSID: urn:lsid:zoobank.org:act:3CED40AC-71DB-4DF0-AFA8-FB56B1862FF2
Material examined. JAPAN, Chichijima Island , Bonin Islands – Ougiura Beach (Holotype, NMW.Z.2022.001.0002, af; imaged paratypes, NMW.Z.2022.001.0003, af, pf; paratypes, NMW.Z.2022.001.0004, 8af, 4f, 4 loose palps; NSMT-Pol P-890, 7af (etoh); imaged paratypes, NMW.Z.2022.001.0005, 3af (formalin fixed); paratypes, NMW. Z.2022.001.0006, 2af, 4f, 2 loose palps (formalin fixed; last chaetiger of 67 chaetiger specimen dissected and slide mounted); NSMT-Pol P-891, 2af (formalin fixed); dissected paratype, NMW.Z.2022.001.0007, af; SEM stub mounted paratypes (dried and sputter-coated), NMW.Z.2022.001.0008–9, 2af), (27.0738, 142.2019), sandy sediment, collected by NJ, by snorkelling, 1–2 m depth, 27/03/2015. GoogleMaps Omura Beach (Paratypes, NMW.Z.2022.001.0010, af; NSMT-Pol P-892, af), (27.0933, 142.1938), sandy sediment, collected by NJ by snorkelling, 1–2 m depth, 26/03/2015. GoogleMaps
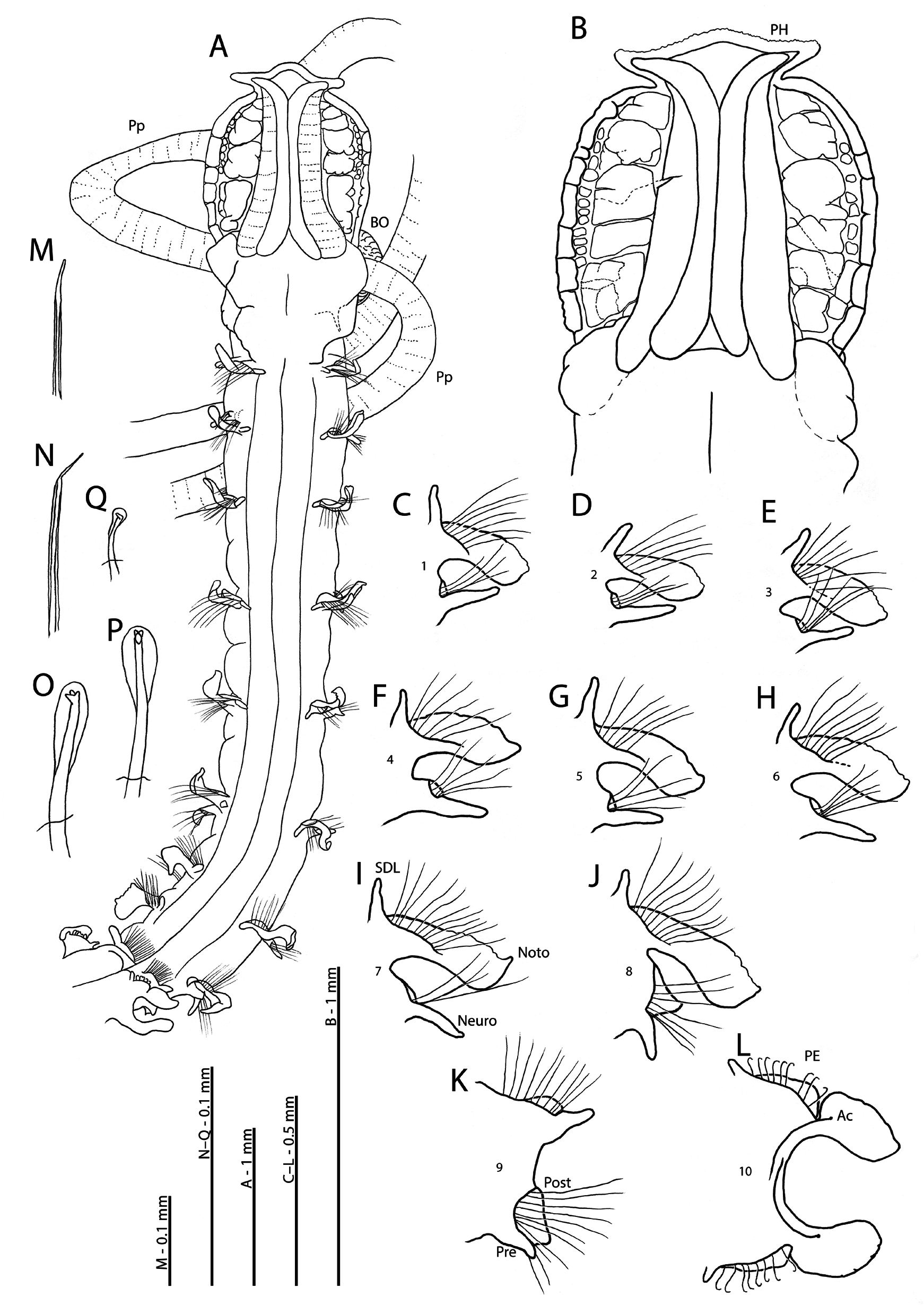
Diagnosis. Prostomium longer than wide with distinct prostomial horns. Notopodia of chaetigers 1–8 with slender foliaceous postchaetal lamellae with crenulate margins, expanded as cirriform superior dorsal lobes. Neuropodia with slender triangular ventral lamellae; chaetiger 8 with additional triangular postchaetal lamellae. Notopodia of chaetiger 9 with rounded postchaetal lamellae confluent with prechaetal ridges and expanded as lateral cirriform lobes. Neuropodia of chaetiger 9 similar to preceding chaetiger. All thoracic chaetae bilimbate capillary. Abdominal lateral lamellae basally constricted with postchaetal expansions. Hooded hooks tridentate, in two groups. Posteriorly open pouches present.
Dimensions. A moderately large species ( Figs 14A View FIGURE 14 ; 15E View FIGURE 15 ; 16D View FIGURE 16 ); junction between thorax and abdomen distinct, abdomen thicker and wider than thorax. Thorax appearing dorsoventrally flattened in comparison to more rounded abdomen ( Figs 15F View FIGURE 15 ; 16B View FIGURE 16 ). All specimens posteriorly incomplete, one posterior fragment present in vial ( Fig. 16F View FIGURE 16 ). Thoracic chaetiger length approximately one and a half times width from chaetiger 3 onwards ( Figs 14A View FIGURE 14 ; 15B View FIGURE 15 ; 16B, C View FIGURE 16 ).
Holotype dimensions: prostomium 1.25 mm long, 1 mm wide; thorax 7.0 mm long (including prostomium), 0.6 mm wide; abdomen 1.0 mm wide; total length approximately 37.0 mm for 65 chaetigers. Posterior fragment 68 chaetigers long. Other paratypes with 17–103 chaetigers for 7.5–44.4 mm.
Description. Prostomium longer than wide (L:W ratio 1.07–1.43); spatulate, anterior margin triangular, appearing smooth, but minute crenulations apparent across entire margin only under high magnification ( Figs 14B View FIGURE 14 ; 18J View FIGURE 18 ). Conspicuous prostomial horns present, separated from distal prostomial margin by a large degree ( Figs 14A, B View FIGURE 14 ; 15A, B View FIGURE 15 ; 16A View FIGURE 16 ; 17D View FIGURE 17 ; 18J View FIGURE 18 ). Lateral margins often appearing undulating due to length of prostomium. Two pairs of prominent longitudinal dorsal muscular ridges present, outer pair ridged transversely and abutting inners for entire length: inner pair diverging distally into each horn. Distinct patterned areas either side of ridges, oblong of varying sizes, additional small circular markings present towards outer edges of the prostomium ( Figs 14B View FIGURE 14 ; 18J View FIGURE 18 ). Burrowing organ almost entirely everted on holotype, heart-shaped, longitudinally ridged inferiorly, upper surface smoother but with some longitudinal ridging ( Fig. 16B View FIGURE 16 ). Burrowing organ additionally everted on 24 specimens: heart-shaped when fully everted (ten specimens) ( Fig. 17B View FIGURE 17 ), round to oval when partially everted (14 specimens) ( Figs 15D View FIGURE 15 ; 16D View FIGURE 16 ).
Right-hand palp retained on holotype, extending to approximately chaetiger 43, non-papillated region reaching mid chaetiger 3. Papillae long, lengthening towards distal palp tips, slender, digitiform ( Figs 15A, B, D, E View FIGURE 15 ; 16A, D View FIGURE 16 ). Papillae in three rows either side of an indistinct longitudinal line proximally and medially, reducing to one row either side at distal tips ( Fig. 17A, C, D View FIGURE 17 ). Palps additionally present (at least partially) on 22 paratypes, long and slender; arising ventrolaterally from base of prostomium, reaching chaetigers 24–50.
Achaetous region behind prostomium approximately one and a half times the size of chaetiger 1 ( Figs 14A View FIGURE 14 ; 15A View FIGURE 15 ). Chaetigers 1–7 similar; parapodia biramous ( Figs 14C–I View FIGURE 14 ; 15F View FIGURE 15 ; 16B View FIGURE 16 ; 18A–G View FIGURE 18 ); notopodia with low triangular prechaetal lamellae confluent with large spatulate to slender foliaceous postchaetal lamellae, upper margins lightly crenulated. Notopodial lamellae marginally longer than neuropodial ones. Long slender, cirriform, prechaetal superior dorsal lobes present. Neuropodial pre- and postchaetal lamellae as low ridges, forming distinct cuff-like structures confluent with ventral triangular lamellae beneath chaetal bundle, of similar size along thorax. Slight postchaetal expansion from approximately chaetiger 5.
Chaetiger 8 parapodia similar to preceding chaetigers ( Figs 14J View FIGURE 14 ; 15F View FIGURE 15 ; 16B View FIGURE 16 ; 18H View FIGURE 18 ), however, neuropodia with additional triangular postchaetal lamellae of similar size to the ventral lamellae.
Notopodial prechaetal lamellae of chaetiger 9 as low ridges confluent with rounded postchaetal lamellae (smaller than those of preceding chaetigers) and inferiorly expanded as small cirriform processes; superior dorsal lobes absent ( Figs 14K View FIGURE 14 ; 15F View FIGURE 15 ; 16B View FIGURE 16 ; 18I View FIGURE 18 ). Neuropodia with triangular postchaetal lamellae confluent with low prechaetal ridges and smaller digitiform prechaetal lamellae. Thoracic chaetae smooth, bilimbate capillaries ( Fig. 14M View FIGURE 14 ), marginally longer in notopodia than neuropodia ( Fig. 14C–I View FIGURE 14 ). Distinctly splayed in posterior thorax ( Fig. 14J, K View FIGURE 14 ). Chaetae of mid rami of chaetiger 9 with slightly irregular blades, tips of which are bent at roughly 45 degrees to shaft, not as marked as the irregular blades noted by Brasil (2003) ( Figs 14N View FIGURE 14 ; 18K View FIGURE 18 ). Paired, reniform ventral swellings present from chaetigers 6–9 ( Fig. 15F View FIGURE 15 ).
Abdominal chaetigers shorter in anterior abdomen but lengthening towards posterior ( Fig. 16A View FIGURE 16 ). Abdominal parapodia with large rounded triangular to spatulate lateral lamellae (edges of lamellae minutely wavy in anterior abdomen) of about equal size in both rami; basally constricted ( Figs 14L View FIGURE 14 ; 15F View FIGURE 15 ; 16E View FIGURE 16 ; 17E, F View FIGURE 17 ). Abdominal lamellae reduce to slender digitiform on extreme posterior chaetigers ( Fig. 16F View FIGURE 16 ). Postchaetal expansion of lateral lamellae behind chaetal rows distinct, triangular ( Figs 14L View FIGURE 14 ; 17E View FIGURE 17 ). Expansion reducing in size towards posterior but still evident until ten chaetigers before pygidium. Triangular processes (DML, VML) present at inner margins of chaetal rows, long, present until approximately ten chaetigers before pygidium.
Abdominal chaetae tridentate hooded hooks ( Figs 14O, P View FIGURE 14 ; 17G View FIGURE 17 ; 18L, M View FIGURE 18 ), approximately ten per ramus in anterior abdomen, in two groups vis-à-vis ( Figs 17H View FIGURE 17 ; 18L View FIGURE 18 ), all of a similar size. Group at outer margin of ramus with approximately twice the number of hooks. Hooks reducing to six or seven per ramus towards posterior. Lamellae of each ramus internally supported by a single curved abdominal support (‘acicula’) chaeta ( Figs 14Q View FIGURE 14 ; 18N, O View FIGURE 18 ), terminating in a tridentate(?) hooded hook. Exact dentition of each chaeta difficult to assess due to size but apical teeth present. Paired posteriorly open pouches ( Figs 16A View FIGURE 16 ; 17F View FIGURE 17 ) present on consecutive segments, starting from around chaetigers 32–50 (observed on 20 specimens, remaining specimens too short to observe pouches). Pouches present for holotype on chaetigers 40–65 on right-hand side of body, and chaetigers 42–65 on left. Pouches appear as simple folds, medially split, present until approximately eight chaetigers from the pygidium (observed from posterior fragment, Fig. 16F View FIGURE 16 ).
Eggs present in ten specimens, approximately 70 μm in diameter (clearly visible from chaetigers 27–43).
Posterior fragment with two long, digitiform pygidial cirri either side of a rounded-triangular pygidium; small, rounded ventrally placed anus ( Fig. 16F View FIGURE 16 ).
Colour. No live specimens observed. Colour of preserved specimens uniformly cream in alcohol with light interparapodial abdominal patches ( Figs 15C, E View FIGURE 15 ; 16D View FIGURE 16 ). Methyl Green staining retained for longer period of time on abdomen (almost as transverse bands, with light speckles as interparapodial patches), faint speckled staining apparent at high magnification on thorax (more apparent in posterior thorax, and on ventral surface) ( Fig. 15B, D, F View FIGURE 15 ). Speckled pigmentation along length of palp papillae, and around their bases.
Habitat. Type specimens found at low tide at Ougiura and Omura beaches, Chichijima Island, Bonin Islands, Japan ( Fig. 1 View FIGURE 1 ) .
Distribution. Magelona boninensis sp. nov. is currently only known from Japan.
Etymology. The specific name refers to the type locality of the Bonin Islands, Japan
Remarks. Nineteen species share morphological similarities with Magelona boninensis sp. nov. in possessing distinct prostomial horns, and in the nature of the thoracic lamellae: Magelona alexandrae Magalhães, BaileyBrock & Watling, 2018 ; M. anuheone ; Magelona cerae Hartman & Reish, 1950 ; Magelona cinthyae Magalhães, Bailey-Brock & Watling, 2018 ; M. cornuta ; M. crenulifrons ; Magelona gemmata Mortimer & Mackie, 2003 ; Magelona lusitanica Mortimer, Gil & Fiege, 2011 ; Magelona marianae Hernández-Alcántara & Solís-Weiss, 2000 ; Magelona montera Mortimer, Cassà, Martin & Gil, 2012 ; M. pacifica , Magelona pulchella Mohammad, 1970 ; Magelona sinbadi Mortimer, Cassà, Martin & Gil, 2012 ; Magelona spinifera Hernández-Alcántara & Solís-Weiss, 2000 ; Magelona tehuanensis Hernández-Alcántara & Solís-Weiss, 2000 ; and Magelona spp. D , G, K, and L of Uebelacker & Jones (1984).
The following species differ from the new species in possessing bidentate and not tridentate hooded hooks of the abdomen: M. pacifica , M. marianae , M. crenulifrons , Magelona spp. D and G, M. cerae , M. pulchella , and M. spinifera . The latter species and Magelona sp. D also differing in possessing spines of the abdominal parapodia. The following species differ from Magelona boninensis sp. nov. in possessing prostomia which are similar in length to width, and thoracic notopodial lamellae which are smooth: M. cornuta , M. cinthyae , M. lusitanica , Magelona sp. L, and M. tehuanensis . The new species shares most similarities with M. pacifica , M. montera , M. gemmata , Magelona spp. G and K, M. sinbadi , M. anuheone , and M. alexandrae in possessing prostomia which are longer than wide with distinct prostomial horns which have a degree of separation from the anterior prostomial margin. As noted above M. pacifica differs in the nature of the abdominal hooks. Magelona montera , from the Western IndoPacific, differs in the shape of the prostomial horns which are laterally rounded. Magelona gemmata and M. sinbadi , also from the Western Indo-Pacific, differ from the new species in possessing smooth notopodial thoracic lamellae. Magelona anuheone , from the Eastern Indo-Pacific, and Magelona sp. K , from the Temperate Northern Atlantic, both also possess smooth notopodial thoracic lamellae, the latter species also differs in the neuropodia of chaetiger 9 which are not triangular as in the new species. Magelona alexandrae , from the Eastern Indo-Pacific, shares many similarities with the new species, however, possesses a smooth prostomial margin and crenulate abdominal lamellae. Additionally, the margins of the notopodial thoracic lamellae are much more distinctly crenulate than in the new species.





| NMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |