
Brachysyllis lagunae ( Hartman, 1961 ) Hartman, 1961
|
publication ID |
https://doi.org/ 10.5281/zenodo.211061 |
|
DOI |
https://doi.org/10.5281/zenodo.6179768 |
|
persistent identifier |
https://treatment.plazi.org/id/8663A32F-D11D-6757-C8BC-FA9F2BB76C50 |
|
treatment provided by |
Plazi |
|
scientific name |
Brachysyllis lagunae ( Hartman, 1961 ) |
| status |
comb. nov. |
Brachysyllis lagunae ( Hartman, 1961) View in CoL , new combination
Figs 13–17 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17
Orseis lagunae Hartman, 1961: 64 -65; 1968: 371–372, figs 1–2. Dioplosyllis broadi Mueller & Fauchald, 1976: 19 View in CoL –22, figs 1–6. Dioplosyllis lagunae Kudenov & Harris 1995: 36 View in CoL –38, fig 1.11.
Material examined: California. Holotype of D. broadi (LACM-AFH Poly 1141); 1 spm, preserved in formaldehyde (SIO-BIC A2260), La Jolla, off La Jolla Beach, 32°50.26’N, 32°50.26’N, 15 m depth, Macrocystis pyrifera holdfasts, SCUBA, colls GWR & Phil Zerofski 15 Oct 2010; 1 spm, preserved in ethanol (SIO-BIC A2254), same collection data; 1 spm, posterior end preserved in ethanol (SIO-BIC A2255), anterior end in formaldehyde (SIO- BIC A2256), same collection data; 1 spm, median segments preserved in ethanol (SIO-BIC A2257), anterior and posterior ends in formaldehyde (SIO-BIC A2258), same collection data; 1 spm, preserved in ethanol (SIO-BIC A2261), same collection data.
Comparative material examined: Brachysyllis japonica Imajima & Hartman, 1964 . Holotype LACM-AHF Poly156. Brachysyllis infuscata (Ehlers, 1901) . Holotype ZMB 6742.
Distribution: Only known from southern California.
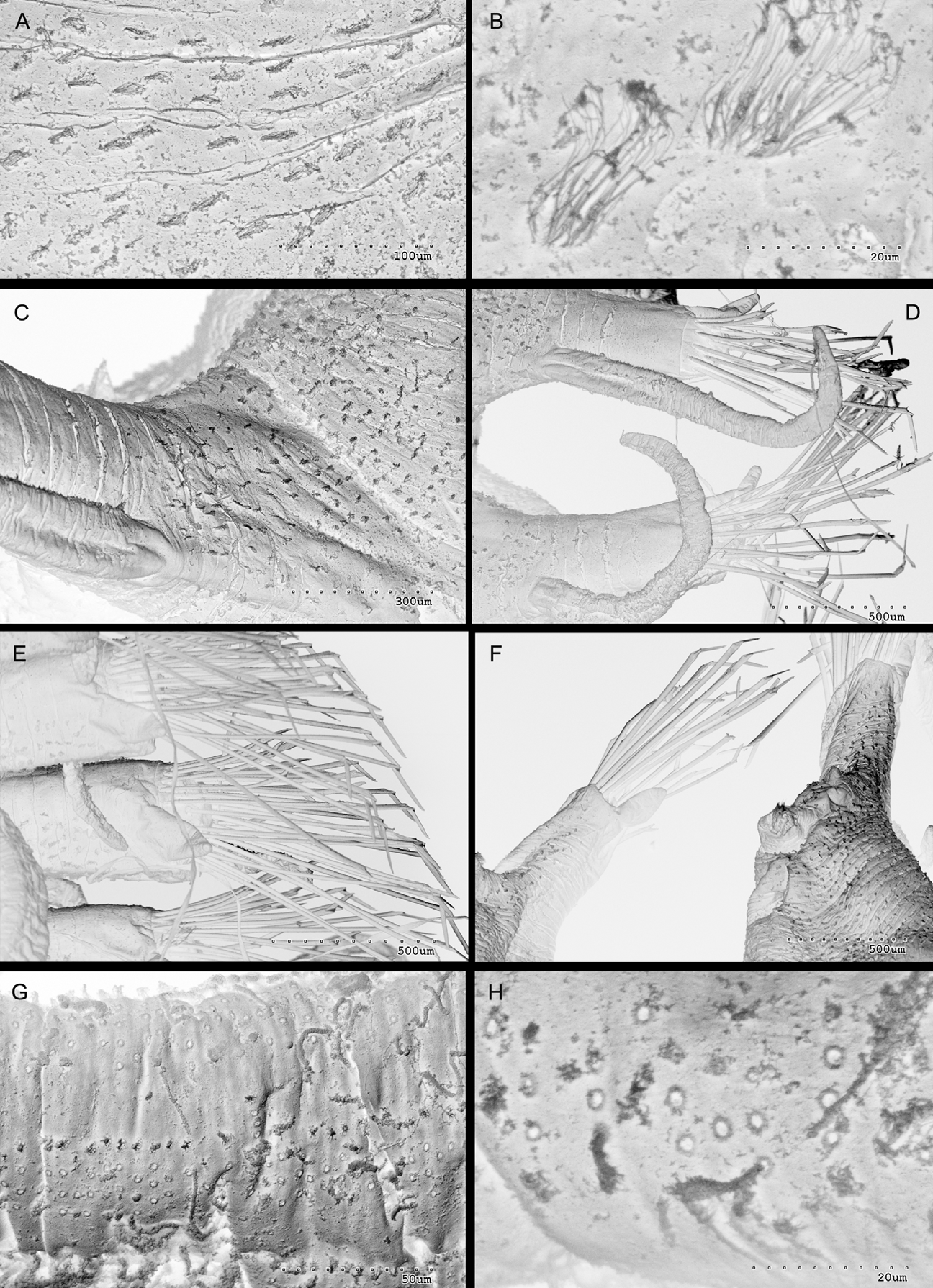
Description: Holotype anterior end 15 mm long for 10 segments, 2.5 mm wide excluding parapodia ( Hartman, 1961). Holotype of D. broady 24 mm long, 2.1 mm wide excluding parapodia, for 13 chaetigers plus one pre-anal achaetous segment ( Mueller & Fauchald, 1976). Longest complete specimen from La Jolla 24 mm long, 2 mm wide excluding parapodia, for 13 chaetigers plus one pre-anal achetous segment ( Fig. 14 View FIGURE 14 F). Body semi-circular in cross-section, dorsum arched; venter flat with longitudinal median groove ( Fig. 14 View FIGURE 14 D). Live specimens with brownish purple dorsal pigmentation ( Figs 13 View FIGURE 13 A, B), venter lighter. Parapodia with white spots on brownish purple ridges. Eyes red. Anterior segments trapezoidal, increasing in size toward posterior end, midbody and posterior segments rectangular ( Fig. 13 View FIGURE 13 A). Segments with series of transverse ridges covered by tufts of cilia, more numerous on venter ( Figs 15 View FIGURE 15 E, F, 16A, B). Parapodia with series of ridges parallel to body axis ( Fig. 16 View FIGURE 16 C), conspicuous on dorsal side ( Fig. 14 View FIGURE 14 D). Prostomium oval, two pairs of eyes in narrow trapezoidal arrangement, eyes large, conspicuous ( Figs 13 View FIGURE 13 A, B). Antennae very long and slender, lateral pair inserted on anterior margin of prostomium, median antenna inserted medially, between anterior pair of eyes ( Figs 13 View FIGURE 13 A, B, 14A, C), short ceratophores present. Palps completely separate, divergent, ventrally folded, twice as long as prostomium, with small lateral papilla on subdistal end ( Fig. 14 View FIGURE 14 C). Paired nuchal organs, densely ciliated, at posterior corners of prostomium ( Fig. 15 View FIGURE 15 A). Triangular areas at both sides of prostomium ( Fig. 14 View FIGURE 14 E). Peristomium shorter than following segments, with two pairs of smooth, long cirri ( Figs. 13 View FIGURE 13 A, B), with short ceratophores, dorsal pair longer than ventral. All dorsal cirri as long as body length ( Figs 13 View FIGURE 13 A, B), smooth to wrinkled, with fine internal granular material, short cirrophores present. Ventral cirri long, digitiform, inserted on proximal half of parapodia ( Figs 14 View FIGURE 14 D, 16D), with internal granular material. Antennae, dorsal and ventral cirri with longitudinal rows of pores, from which cord shaped secretions arise ( Fig. 16 View FIGURE 16 G, H). Parapodial lobes long, more than half midbody segment width, quadrangular with triangular dorsal postchaetal lobes ( Figs 16 View FIGURE 16 E, F). Parapodia with 20–30 compound, heterogomph chaetae, blades distinctly bidentate, with short spines on cutting edge of blades ( Fig. 17 View FIGURE 17 A–F). Blades within facicles decreasing in length from dorsal (140 µm) to ventral (83 µm) ( Fig. 17 View FIGURE 17 C–F). Distal part of fangs with two tips ( Fig. 17 View FIGURE 17 C–F). Five straight aciculae per parapodium, some ending in thin, distally curved tips. Pygidium short ( Fig. 14 View FIGURE 14 F), with long and cirriform paired anal cirri. Pharynx shape not visible by transparency. Several crowns of structures on anterior part of the everted pharynx; a crown of ten distal papillae and a crown of large cilia and five internal conical teeth ( Figs 14 View FIGURE 14 B, 15B–D). Pharyngeal tooth large and conical, located dorsally on anterior inner pharynx surface ( Fig. 15 View FIGURE 15 B, C). Proventricle extending through segment 3–6, with about 90 muscle cell rows (Kudenov & Harris, 1995). Reproduction by epigamy ( D. broadi holotype is an epitokous form).
Habitat: Intertidal and shallow subtidal rocky substrata and holdfasts of Macrocystis pyrifera .
Remarks: The genus Brachysyllis was described by Imajima & Hartman (1964), with B. japonica as the type species and the additional species Amblyosyllis infuscata Ehlers, 1901 as a new combination. Later, Brachysyllis was synonymized with Dioplosyllis Gidholm, 1962 by Imajima (1966). Aguado & San Martín (2008) re-examined the type material of B. infuscata and concluded that the genus was valid and separate from Dioplosyllis . They proposed that three species should be included in the genus: B. japonica , B. infuscata and D. lagunae . Here we confirm that D. lagunae belongs to Brachysyllis . The three species share the diagnostic characters proposed for the genus: body long with a fixed number of chaetigers (13 in all known species) together with one achaetous, pre-anal segment; anterior segments trapezoidal is shape; palps totally free at bases, with a subdistal papilla; ventral cirri long, filiform and proximally located on parapodia; about five distally acute and curved aciculae ( Imajima & Hartman 1964; Mueller & Fauchald 1976; Aguado & San Martín 2008); distribution restricted to the Pacific Ocean ( B. infuscata in Chile; B. japonica in Japan; and B. lagunae in California).
Examination of the type material of B. infuscata , B. japonica and D. broadi (synonimized with B. lagunae by Kudenov & Harris 1995) did not provide distinct morphological differences between the three species. Brachysyllis infuscata and B. lagunae are very similar to each other and only differ from B. japonica in the number of distal teeth on the pharynx (five for the former, six for the latter). However, we were not able to confirm the number of teeth when re-examining the types of B. japonica , since a detailed study of the pharynx may damage it. Although there are no obvious and consistent morphological differences between these three species, the geographical distributions at both sides of the Pacific at present justifies treating B. lagunae as a distinct species. In this study, SEM provides more details about morphological traits of this intriguing genus, such as the presence of series of clumps of cilia, secretion of material from longitudinal rows of pores on dorsal and ventralcirri, and details of the pharyngeal structures. Hartman (1961) described B. lagunae as a hesionid of the genus Orseis probably because of the large size of these animals and their remarkable morphology, which make them appear superficially quite different to the rest of syllids. Phylogenetic analyses of Syllidae based on morphology ( Aguado & San Martín 2009) revealed the genus Brachysyllis as sister to Amblyosyllis Grube, 1857 , and both together with Dioplosyllis . The three genera were supported only by the presence of very long lateral antennae and dorsal cirri. In contrast, a clade including Amblyosyllis and Brachysyllis was supported by four apomorphies: macrofaunal body size with few segments, constant number of segments, anterior segments trapezoidal in shape, and presence of a pre-anal segment without chaetae. Additionally, series of clumps of cilia and pores on cirri with similar secretions have been seen in some Amblyosyllis species (pers. obs.).
The sequence of COI (GenBank accession number JQ623498 View Materials ) of Brachysyllis lagunae showed a BLAST identity of ~ 78% to a range of polychaetes; none of these are syllids, but clearly further sequencing of other genes will be needed to resolve its systematic position.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Brachysyllis lagunae ( Hartman, 1961 )
| Pleijel, Fredrik, Aguado, Maria Teresa & Rouse, Greg W. 2012 |
Orseis lagunae
| Mueller 1976: 19 |
| Hartman 1961: 64 |