
Gianius navarroi, Rodriguez, Pilar & Achurra, Ainara, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.193114 |
|
DOI |
https://doi.org/10.5281/zenodo.6211155 |
|
persistent identifier |
https://treatment.plazi.org/id/C8764570-FFA6-AA0A-71C8-BED3FE93FB4F |
|
treatment provided by |
Plazi |
|
scientific name |
Gianius navarroi |
| status |
sp. nov. |
Gianius navarroi n. sp.
(Figures 1–2)
Phallodrilinae n. sp. in Camacho et al. 2006.
Holotype. MNCN 16.03/3051. An incomplete mature specimen, stained in Ehrlich’s Haematoxylin and whole-mounted in Canada balsam.
Type locality. Ojo Guareña cave (site: “Granja Ortiz”, 1 June 2002), northern Burgos, Spain. UTM coordinates for the Palomera entrance: X:446595, Y:4764790, Z:724.
Paratypes. MNCN 16.03/3052. A complete mature specimen (unmated) from the type locality, site: “Gour de las Hojas” (8 March 2003). A complete mature specimen in the collection at the University of the Basque Country (UPV/EHU), from the type locality, 11 June 2007. Both specimens are stained in Ehrlich’s Haematoxylin and whole-mounted in Canada balsam.
Etymology. Named after the physiologist Dr. Enrique Navarro who has helped P. Rodriguez in sampling numerous sites for the study of aquatic oligochaetes, and has collaborated in applied research with oligochaetes.
Description. Length of preserved worms 1.7 mm; number of segments (in two complete specimens) 23 and 26; maximum diameter 150 µm in segment XI. Prostomium rounded, with glandular epidermis (Fig. 1A). Pygidium short and wide (Fig. 1B). Body wall consisting of a fine cuticule (ca 1 µm thick), an epidermal layer (5 µm thick) and a relatively poorly developed musculature (2–3 µm thick). Clitellum formed by glands disposed in one layer, from the last third of segment X back to the beginning of segment XII. There is no secondary annulation. One pair of simple spermathecal pores in the most anterior part of segment X, opening in line with ventral chaetae. One pair of male pores in segment XI on porophores, in line with ventral chaetae. All chaetae bifid, 24–27 µm long, with distal tooth shorter than proximal. Anterior segments with 2–5 chaetae per bundle, 2 in postclitellar segments. Chaetal bundles are in the posterior third of the segment. A bundle of 5 penial chaetae forms an arc, towards the sagittal plane of the body ( Fig. 2 View FIGURE 2 ). Penial chaetae straight, with bent distal end, bifid, and of similar length to somatic chaetae.
Gianius navarroi A: anterior part of the body in dorso-ventral view, IV–VII: segment number; B: anterior and posterior ends of the body in lateral view; C: segments VII and VIII. Abbreviations: b: brain, cc: chloragogen cells, i: intestine, od: oesophageal diverticula, ph: pharynx, phg: pharyngeal glands, py: pygidium. Scale bars: A and B: 100 µm, C: 50 µm.
Brain with two histologically distinct sections: anterior and posterior parts having large nuclei intensely stained with haematoxylin, and a granulated median part that does not stain well. No coelomocytes observed within the coelomic cavity. Pharyngeal glands in segments III–V, dorsal in III, and both lateral and ventral in segments IV and V (in a paratype, glands also ventrally in VI). Chloragogen cells form a layer on the gut from segment VI backwards. In segment VII, a pair of dorsal digestive diverticula, can occupy half or the entire length of the segment, and appear to be attached to the oesophagus at the posterior end of the segment (Figs. 1A, C). A dilatation of the gut in segment VIII is interpreted as the beginning of the intestine (Figs. 1B, C).
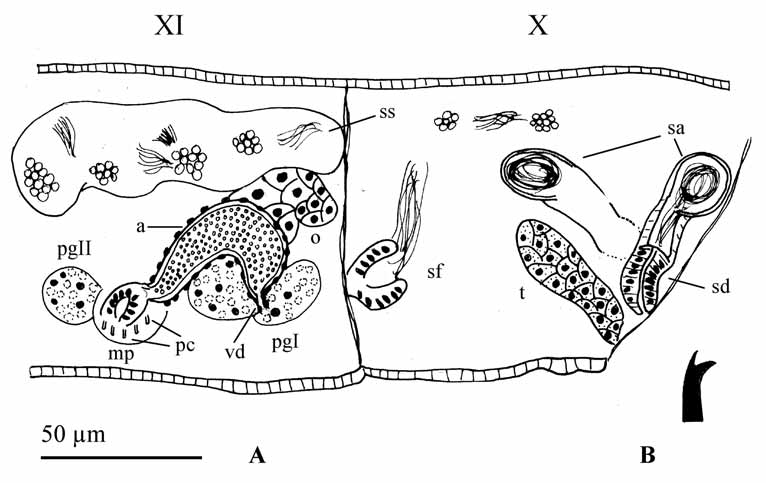
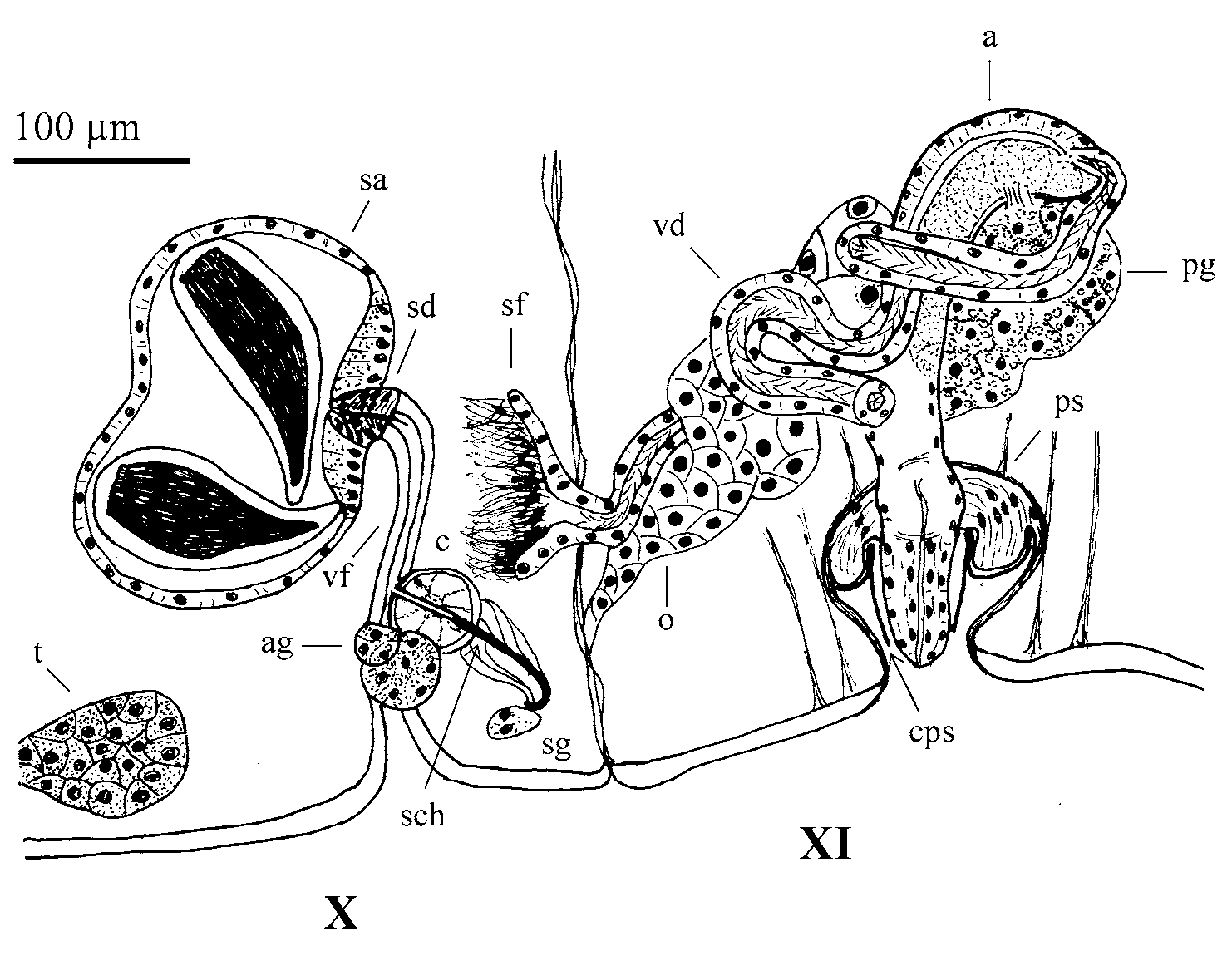
One pair testes in segment X and one pair ovaries in segment XI. Sperm sacs in segment XI (in one specimen spermatogonia and spermatocytes are free in the coelom of segment X). Male ducts paired, with two prostate glands: one ental, close to the junction of the vas deferens with atrium; and other ectal, close to the male pore, much smaller than the ental one. A ciliated vas deferens (6 µm diameter) joins the atrium apically. Atria tubular (77 µm long, up to 15 µm diameter) and curved; thicker in the middle in lateral view (with homogeneous diameter in dorsal view) and gradually narrowing to both ental and ectal ends. The atrial musculature is thin (2–3 µm thick) and the lumen is ciliated or filled with granules (as in Fig. 2 View FIGURE 2 ). The atria open into salient round porophores (ca 10 µm long), where the penial chaetae are located. Female funnels were not observed.
One pair of spermathecae, with a stout duct (20 µm long, 13–16 µm wide) and an elongated globular ampulla (40–50 µm long, 16–17 µm maximum diameter), which contains sperm forming a loop.
Distribution and habitat. Gianius navarroi n. sp. is known only from Ojo Guareña cave system, and it is likely to be a stygobiont. The species was collected from two different epikarstic sites within the cave, but was rare, at most one specimen per sample. Habitats sampled were puddles and cave streams, having temporary or permanent water, at depths up to 40 cm; sediment was composed of mud, sand and/or stones. Water temperature was 8.4–9.9ºC, pH 7.8–8.0, dissolved oxygen 74.3–78.4 % saturation and conductivity 389–465 µS/cm.
Taxonomic remarks. Gianius navarroi n. sp. has spermathecal pores in line with the ventral chaetae, spermathecae with short ducts, vas deferens entering the apical end of an elongated and curved atrium, and two prostate glands per atrium attached to proximal and distal ends, respectively. All of these characteristics are diagnostic for the phallodriline genus Gianius Erséus, 1992 . The presence of digestive diverticula in segment VII was previously known only in the species Gianius labouichensis ( Rodriguez and Giani, 1989) ; although the connection of the diverticula with the gut appears to be in the posterior end of the segment for the new species, whereas for G. labouchensis the single diverticulum connects anteriorly. However, the latter is a distinctive species, well distinguished from other congeners by the lack of penial chaetae, a uniformly tubular atrium with a protrusible pseudopenis, and elongated narrow spermathecal ampulla. The general structure of the male duct in G. navarroi n. sp. is more similar to the remaining species of the genus, being closest to G. aquaedulcis ( Hrabë, 1960) , G. cavealis Juget and des Chatelliers, 2001 and G. anatolicus Arslan, Timm and Erséus, 2007 based on the thin atrial musculature and the moderate (4–5) number of penial chaetae. Gianius cavealis is well distinguished from the new species by its distal atrial duct well separated from an atrial ampulla, and G. anatolicus by the presence of a long, simple spermathecal duct and three different types of somatic chaetae. The general structure of male duct and spermatheca are similar to G. aquaedulcis , and differences are mainly related to the relative size of the prostate glands (both glands large and of about equal size, in G. aquaedulcis but of visibly different size in the new species), as well as the size of chaetae (41 µm in G. aquaedulcis ) and the presence of oesophageal digestive diverticula. The G. aquaedulcis material described by Farara and Erséus (1991) in North America has a larger body size and wider atria (33–38 µm width); it is distinguished from the new species by the absence of noticeable porophores at the male pores, (described as “simple male pore on inconspicuous papilla”), and by the blunt ental end of the atria. The shape of the pygidium in G. n a v a r ro i is unusual, but very similar to that described for Mexidrilus obtusus by Erséus (1992: Fig.13).
Gianius navarroi n. sp. is separated from the previously known species of the genus by the presence of oesophageal diverticula, salient round porophores with the penial chaetae, and marked difference in size of prostate glands (ental gland larger than distal).
Isochaetides gianii n. sp. ( Figures 3–5 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Lamadrilus sp. in Achurra and Rodriguez (2008)
Holotype. MNCN 16.03/3053. A dissected worm, stained in Ehrlich’s Haematoxylin and mounted in Canada balsam.
Type locality. Apraiz karstic spring (6 September 2005), Santa Eufemia–Ereñozar karst unit, Biscay, Spain. UTM coordinates X:524225, Y:4801350, Z: 22.
Paratypes. USNM 2052660. A dissected specimen from the type locality (6 September 2005). MNCN 16.03/3054: two whole-mounted specimens from Argatxa spring (30 September 2005). In the collection at the University of the Basque Country (UPV/EHU): a sectioned and a dissected specimen from the type locality (6 September 2005); a dissected specimen from Oxiña spring (1 February 1985). All sites in Santa Eufemia– Ereñozar karstic unit, Biscay, Spain.
Other material. Three specimens preserved in 70% alcohol from the type locality (6 September 2005). Two dissected and three specimens in 70% alcohol, from Argatxa spring (30 September 2005). Four sectioned, seven dissected, two whole-mounted and 17 specimens preserved in 70% alcohol, from Aulestia spring (23 February 1985). All in the collection at the University of the Basque Country (UPV/EHU).
Other localities. Argatxa spring: UTM coordinates X:527790, Y:4800925, Z:2; Oxiña spring: X:529825, Y:4801300, Z:38; Aulestia spring: X: 536494, Y: 4794430, Z:66. All in Santa Eufemia–Ereñozar karst unit.
Etymology. Named after Dr. Narcisse Giani for his important contribution to the taxonomy and general biology of aquatic oligochaetes and also for his important role in the formation of many oligochaete taxonomists (including the first author).
Description. Body length 8–10 mm; segment number 59–83; mean body diameter 260 µm (range: 180– 335 µm) in segment V, and 324 µm (225–390 µm) in segment VIII. Prostomium rounded or conical, 75–125 µm long, width about the same or wider at its base (all measurements in fixed individuals). Body wall consisting of a thin cuticle (<1 µm), smooth epidermis (5–10 µm thick, but up to 22 µm in the clitellum) and relatively poorly developed musculature (5–7 µm thick). There is no secondary annulation. Clitellum formed by a single layer of glandular cells, extended from anterior part of XI, back to the line of chaetae in segment XII; no glands in the region between the male pores. One pair of deep folds hiding spermathecal apertures (irregular in shape) in segment X, in line with ventral chaetae. One pair of male pores in segment XI, close to and slightly lateral to the ventral chaetae. In some specimens, the ventral part of segment XI appears deeply retracted by the action of dorso-ventral muscle strands.
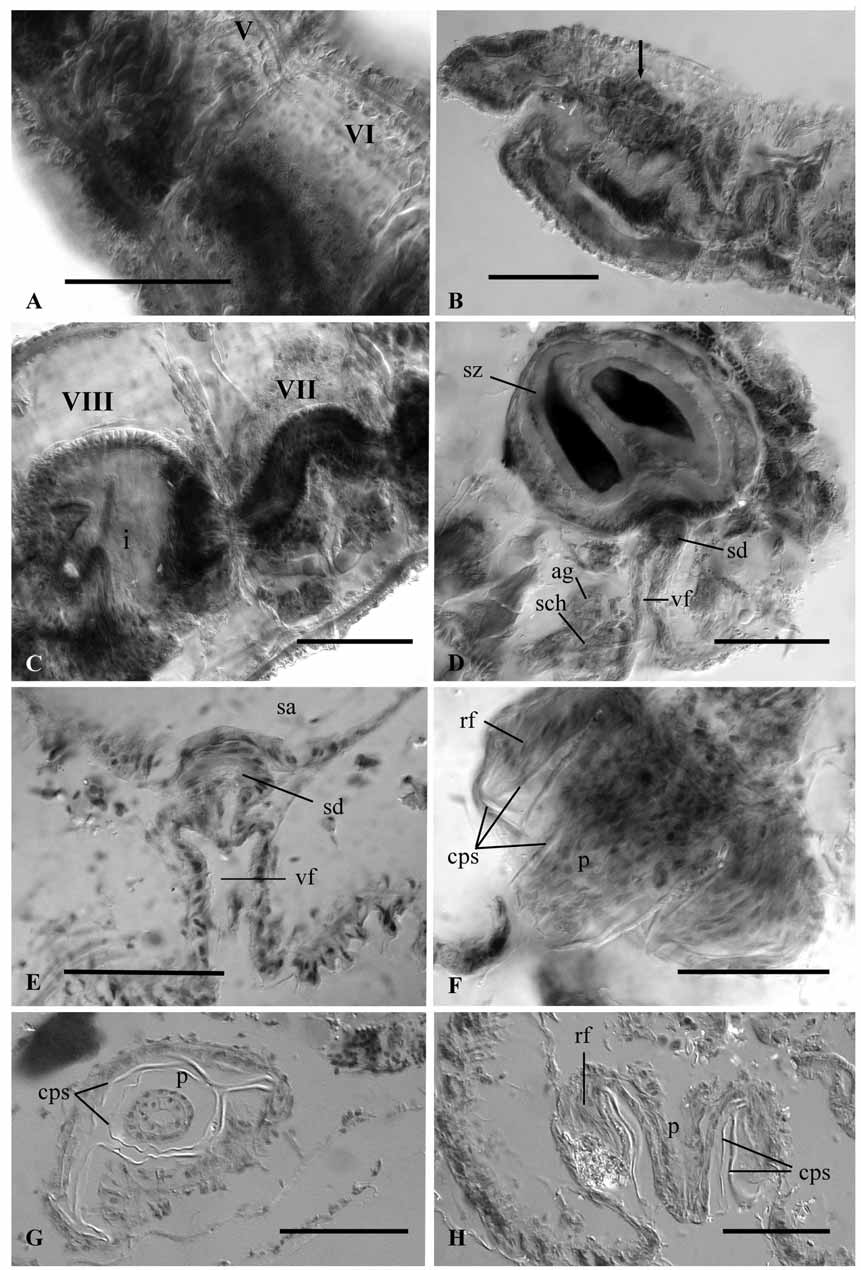
Ventral chaetae 4–6 per bundle, reduced to two in posterior part of the body, 50–75 μm long, with distal tooth thinner and shorter than proximal ( Fig. 3 View FIGURE 3 A, B). Hair and pectinate chaetae in dorsal bundles ( Fig. 3 View FIGURE 3 C–E). Hair chaetae in preclitellar region 1–3 (seldom 4) per bundle, 162–262 µm long; in postclitellar region 1–2 per bundle, 75–125 µm long, gradually becoming shorter backwards. Pectinate chaetae in preclitellar segments 2– 4 per bundle, 47–90 µm long and with distal nodulus (at about ⅓ to ¼ from the distal end); 2–3 somewhat shorter chaetae, posteriorly. Teeth of pectinate chaetae about equally long in anterior segments, but distal tooth shorter and thinner than proximal in posterior segments. The number of intermediate teeth in pectinate chaetae 1 or (usually) 2 in anterior dorsal chaetae; intermediate teeth are absent in posterior segments.
Modified, distally grooved spermathecal chaetae in segment X, one per bundle, 102–145 µm long and 2– 3 µm thick ( Fig. 3 View FIGURE 3 F). There are no modified penial chaetae; 1–2 somatic ventral chaetae per bundle, 50–52 µm long, are found ventrally in segment XI, lateral to the male pores.
Brain extends back to 1/2. Coelomocytes absent or, if present, they are scarce, small, oval (5 µm maximum diameter), nucleated and granular. Commissural blood vessels connecting dorsal and ventral vessels are visible in most examined specimens; they form several loops in anterior segments, particularly in V ( Fig. 4 View FIGURE 4 A). Typical oligochaete dorsal, muscular pharyngeal pad not developed. In segments II–III, pharynx with well-developed epithelium both dorsal and ventrally, and a large mass of glands dorsally ( Fig. 4 View FIGURE 4 B). A few clusters of pharingeal glands are located dorso-laterally to the digestive tract in segment IV and laterally in V. A layer of chloragogen cells on the gut extends from segment VI backwards. In segment VIII, oesophagus opens abruptly into the intestine ( Fig. 4 View FIGURE 4 C).
One pair of spermathecae in segment X. A short spermathecal duct (30–50 µm long, 30–50 µm wide), with a thick epithelium, opens ventrally in segment X at the inner end of a deep epidermal fold ( Fig. 4 View FIGURE 4 D, E). Spherical to globular spermathecal ampulla (175–220 µm maximum diameter), with a thin epithelium (3–7 µm high, although up to 20 µm close to the connection with the spermathecal duct). The ampulla usually contains 4–5 drop-like, elongate spermatozeugmata (125–202 µm long, 35–82 µm maximum diameter), which consist of an external clear matrix 7–15 µm thick around an axial cylinder of longitudinally arranged fertilizing sperm ( Fig. 4 View FIGURE 4 D). The spermathecal chaetae are orientated with their distal ends towards the spermathecal fold. The proximal end of the chaeta is bent, and associated with a chaetal gland (follicular cells) with several muscular strands; the distal end (ca one third of its total length) is grooved and contained within a chaetophore (32–50 µm diameter), which is internally glandular, radially arranged and externally surrounded by a muscular wall about 2–3 µm thick ( Fig. 5 View FIGURE 5 ). Close to the chaetophore, and most likely functionally associated with it, 1–3 accessory glands can be observed, with histological structure similar to that of the prostate gland. The position of the chaetophore is usually anterior to the spermathecal fold, although in some cases it can be posterior to the fold.
One pair testes in X and one pair ovaries in XI. Sperm sacs may extend to IX anteriorly and to XII posteriorly. Egg sac can reach segment XIII. Sperm funnel 80–127 µm high in lateral view (about one third the diameter of the body), attached to septum 10/11. Vas deferens ciliated throughout, two to three times longer than the atrium and penis, and restricted to segment XI, where it forms a few loops. It is up to 30 µm in diameter, but thinner in its proximal end (down to 13 µm diameter), and in some specimens also thinner just before its junction with the atrium (15 µm diameter). Atria in XI, tubular (250–300 µm long), bent and expanded near proximal end (42–60 µm, measured at the level of junction with the prostate), thinner in their distal part (30–40 µm), and swelling before entering the penial sac ( Fig. 5 View FIGURE 5 ). Distal section of atrium may be bent, forming a loop before entering the penial sac. Two different types of atrial cells, one forming a dense layer (up to 37 µm high) at the concave side of the atrium, and the other type with a clear granulated appearance forming a layer of cells (up of 20 µm high) along the convex side and the proximal part of the atrium. Atrial musculature very thin (<2 µm thick) and atrial lumen narrow, 2–5 µm. A medium-size, compact prostate gland connects by a short stalk to the concave side, at the proximal expanded end of the atrium. Vas deferens enters subapically on the convex side of the atrium. Penial sac well developed, oval, with maximum diameter parallel to the longitudinal axis of the body (115–145 µm), having a ring fold at its inner end. Penis cylindrical (65–115 µm long and 35–50 µm diameter) and formed by two epithelial layers ( Fig. 4 View FIGURE 4 F–H). A mushroom-shaped cuticular layer, consisting of an inner oval part (2–4 µm thick) lining both the penial sac and the proximal ring fold; and a central stalk (1–2 µm thick) forming a cuticular sheath around the penis. This penial sheath is slightly conical and encloses most of the penis, which is free inside it and can project partially outwards. Female funnels were not observed.
Teratology. One specimen with two male ducts plus one spermatheca on one side, and one male duct, one spermatheca plus two chaetophores on the other side.
Distribution and habitat. The new species was found in four karstic springs located in the karst of Santa Eufemia-Ereñozar, in sandy sediments. Water characteristics were for Argatxa spring: 13.7ºC, pH 7.2, 446 µS/ cm and 91 % dissolved oxygen; and for Apraiz spring: 14.8ºC, pH 7.0, 469 µS/cm and 70 % dissolved oxygen.
Taxonomic remarks. The position and general structure of the gonads, male ducts and spermathecae in the new species are characteristic of the oligochaetes of the subfamily Tubificinae . The new species is related to the tubificine genera Isochaetides Hrabë, 1966 and Lamadrilus Timm, 1998 by the presence of a tubular atrium, a discrete prostate gland attached to atrium by a narrow stalk, a vas deferens longer than the atrium and ciliated throughout, a penis within a penial sac, and modified spermathecal chaetae. Other close genera with similar characteristics of the male duct are Tasserkidrilus , Arctodrilus , Troglodrilus and Haber , the first three lacking modified genital setae, and the latter with both spermathecal and penial chaetae modified. The new species has modified spermathecal setae in X and no penial chaetae, as well as a general structure of the male duct which can be present in either Isochaetides or Lamadrilus. These include the vas deferens joining the atrium subapically, and the expanded proximal end of the atrium, which has two layers of glandular cells: a dark, densely granulated layer in the concave side and a light layer in the convex side. The new species is well distinguished from other species of Isochaetides or Lamadrilus by a combination of characters that include: hair chaetae and pectinates in dorsal bundles, grooved spermathecal chaetae associated with chaetophores and glands, a well-developed penial sac lined by a cuticle layer with a proximal ring fold, and a cuticular penial sheath (not asymmetrical or in the form of a spade).
Due to the feeble differences between the genera Isochaetides and Lamadrilus, the new species was first identified as Lamadrilus sp. ( Achurra & Rodriguez 2008), mainly based on the presence of hair chaetae. Most species in this genus have only bifid somatic chaetae, except L. bazikalovae ( Chekanovskaya, 1975) , which presents different rates of reduction in hair chaetae and pectinates ( Timm 1998). However, we agree with Brinkhurst and Kathman (1983) that the absence of species with hair chaetae in a genus should not be a problem for the inclusion of a new species with hair chaetae; and I. gianii n. sp. has been classified on the basis of the structure of the reproductive organs. The genus Isochaetides Hrabë, 1966 includes species that possess modified spermathecal chaetae (except I. distinctus Semernoy, 2001 ). These are characteristically grooved in the distal end (except for I. michaelseni Lastočkin, 1937 , which has bifid genital chaetae), and generally appear associated with spermathecal glands. In several species, these glands are poorly described, but their remarkable distal position with respect to the spermathecal chaeta, and their structure (sometimes only seen in drawings), is similar to the chaetophores that we have described in I. gianii n. sp. Chaetophores associated with spermathecal chaetae are also known in other oligochaete genera [e.g. the rhyacodriline Protuberodrilus tourenqui Giani & Martínez-Ansemil, 1979 and the telmatodriline Alexandrovia ringulata ( Sokolskaya, 1961) ]. On the other hand, only one of the six known species of Lamadrilus ( L. sorosi Timm, 1998 ) has modified spermathecal chaetae of the same type as that described in Isochaetides species, as well as a chaetophore and accessory glands associated to the chaetal sac.
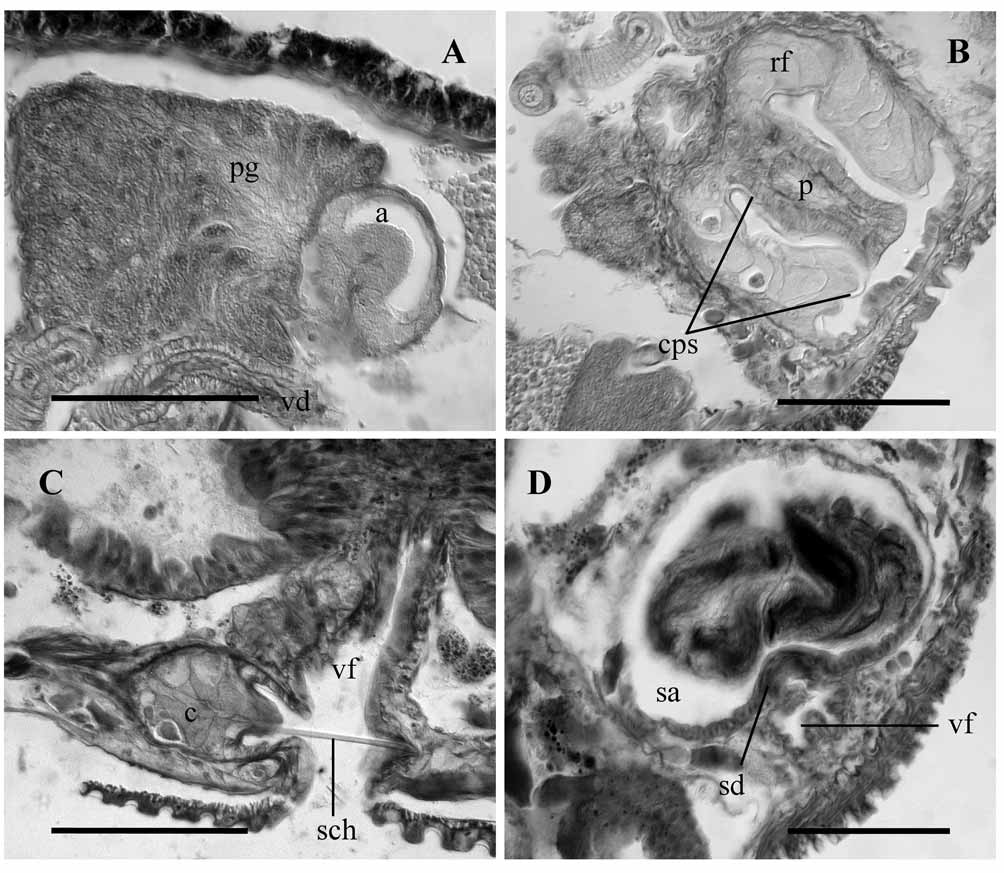
The two different epithelial layers of the atrium described in the new species are probably equivalent to the dark and light layers described in L. sorosi by Timm (1998), and are also reminiscent of the layers shown in drawings of some Isochaetides species. This histological characteristic has been confirmed in the examination of type specimens of I. freyi ( Brinkhurst, 1965) ( Fig. 6 View FIGURE 6 A), although it has probably been overlooked in most descriptions, not only in species of these genera but also in other tubificines.
The presence of a penial sac with an apical ring fold is known in other Isochaetides species (e.g. I. freyi ) ( Fig. 6 View FIGURE 6 B). Several lateral folds are present in the penial sac of some Lamadrilus species (e.g. L. sorosi ), but never in the form of an ental ring fold. The presence of a short, mostly asymmetrical cuticular penial sheath (sometimes with a spade-shaped distal appendage) is a diagnostic character in Lamadrilus; however, the genus incorporates different types of penial sheaths (sometimes variable in shape within the same species, with a more or less marked asymmetry, see Timm, 1998: Figs. 45–50). The diagnosis of the genus Isochaetides by Snimschikova (1998) describes the cuticular penial sheath as thin or absent and Semernoy (2004) describes it as more or less thin, either of uniform width or conical but without oblique cut on its distal end. In fact, all Isochaetides species known to date, except for I. lacustris , have a thin cuticular penial sheath or a thin cuticular covering on the penis (although thick in I. distinctus Semernoy, 2004 ). The examination of the type specimens of I. freyi have revealed a conspicuous cuticular layer, covering the inner side of the penial sac, the large ental fold and the penis, and taking the shape of their contour ( Fig. 6 View FIGURE 6 B), in a similar way to that in the new species.
The structure of the spermatheca is not commonly used as a diagnostic character in tubificines. However, in Isochaetides the short spermathecal ducts, associated with modified spermathecal chaetae located in adjacent glandular sacs (referred to as chaetophores in present work), appear to be a very common feature. The structure of the spermathecal duct is contentious and what is described here as a short spermathecal duct opening at the ental end of a ventral body fold, appears to be interpreted as a short ental sphincter of the spermathecal duct by Semernoy (2004) in I. peniacerus Semernoy, 2004 , I. compactus Semernoy, 2004 and I. septatus Semernoy, 2004 . The histology of the duct in the new species reveals that the cells are not muscular, and the irregular aperture at the body surface does not have the structure of a pore, typically formed by cells arranged in a circle around the opening. Short spermathecal ducts, opening into ventral folds of the body wall, associated with grooved spermathecal chaetae and chaetophores, are also present in I. freyi ( Fig. 6 View FIGURE 6 C, D). These characters are shared by most Isochaetides species, and we have considered them together with the general structure of the male duct and penis, for the classification of the new species as a member of the genus Isochaetides .
The finding of an Isochaetides species in karstic areas in northern Iberian Peninsula is noteworthy, since the genus is represented mainly in eastern Europe (Timm & Veldhuijzen van Zanten 2002) and Asia (many species in Lake Baikal, see Semernoy 2004), although three species have been described in America: I. freyi ( Brinkhurst, 1965) and I. curvisetosus ( Brinkhurst & Cook, 1966) [considered as a probably synonym of I. hamatus ( Moore, 1905) after Brinkhurst, 1986] in North America; and I. lacustris (Černosvitov, 1939) in South America.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |