
Mesobiotus romani, Roszkowska & Stec & Gawlak & Kaczmarek, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4450.5.2 |
|
publication LSID |
lsid:zoobank.org:pub:E6BBACAA-2683-4443-A66A-86D848CF4C5C |
|
DOI |
https://doi.org/10.5281/zenodo.5978219 |
|
persistent identifier |
https://treatment.plazi.org/id/48773A43-240E-164C-FF15-FB57FBAF2808 |
|
treatment provided by |
Plazi |
|
scientific name |
Mesobiotus romani |
| status |
sp. nov. |
Mesobiotus romani View in CoL sp. nov.
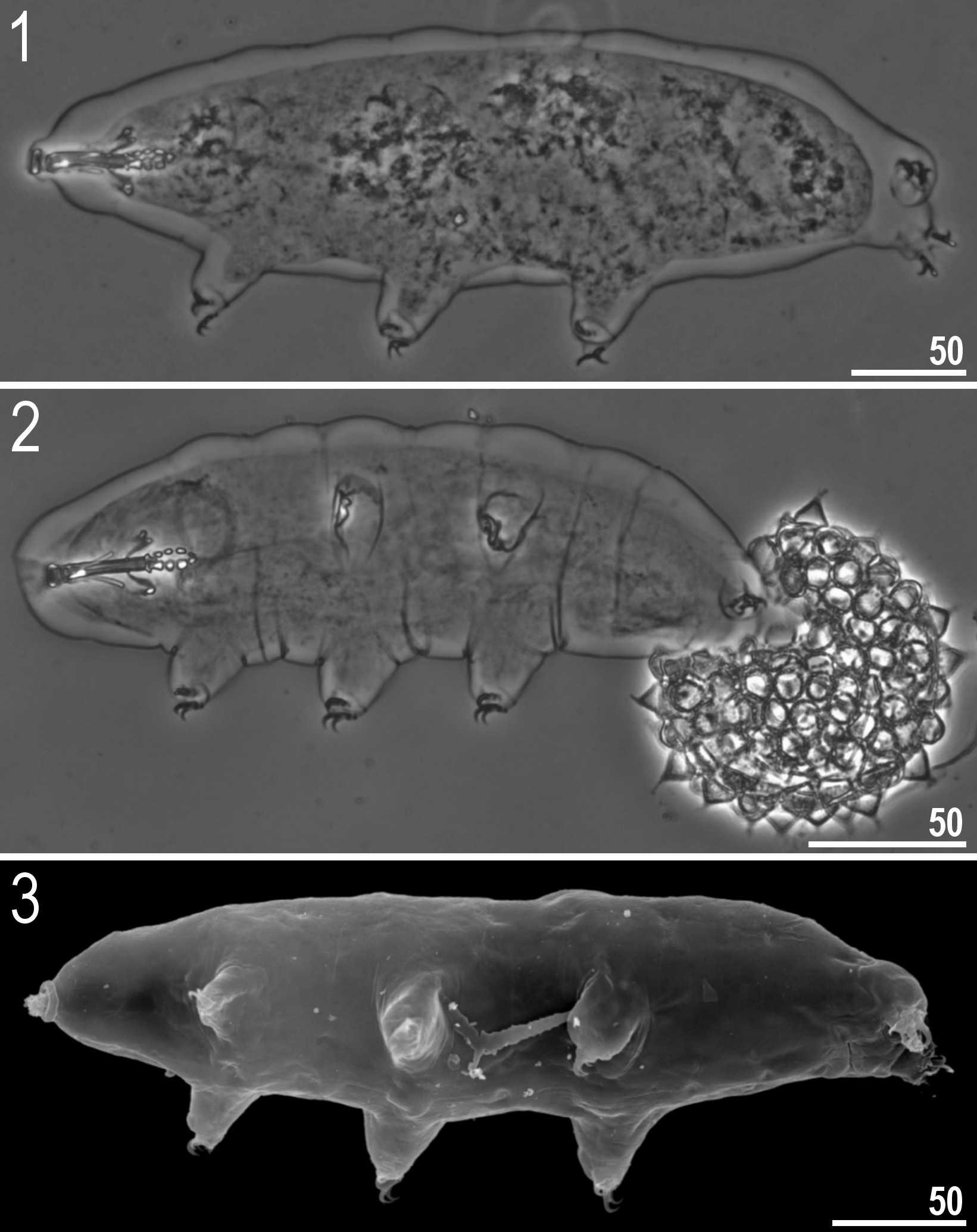
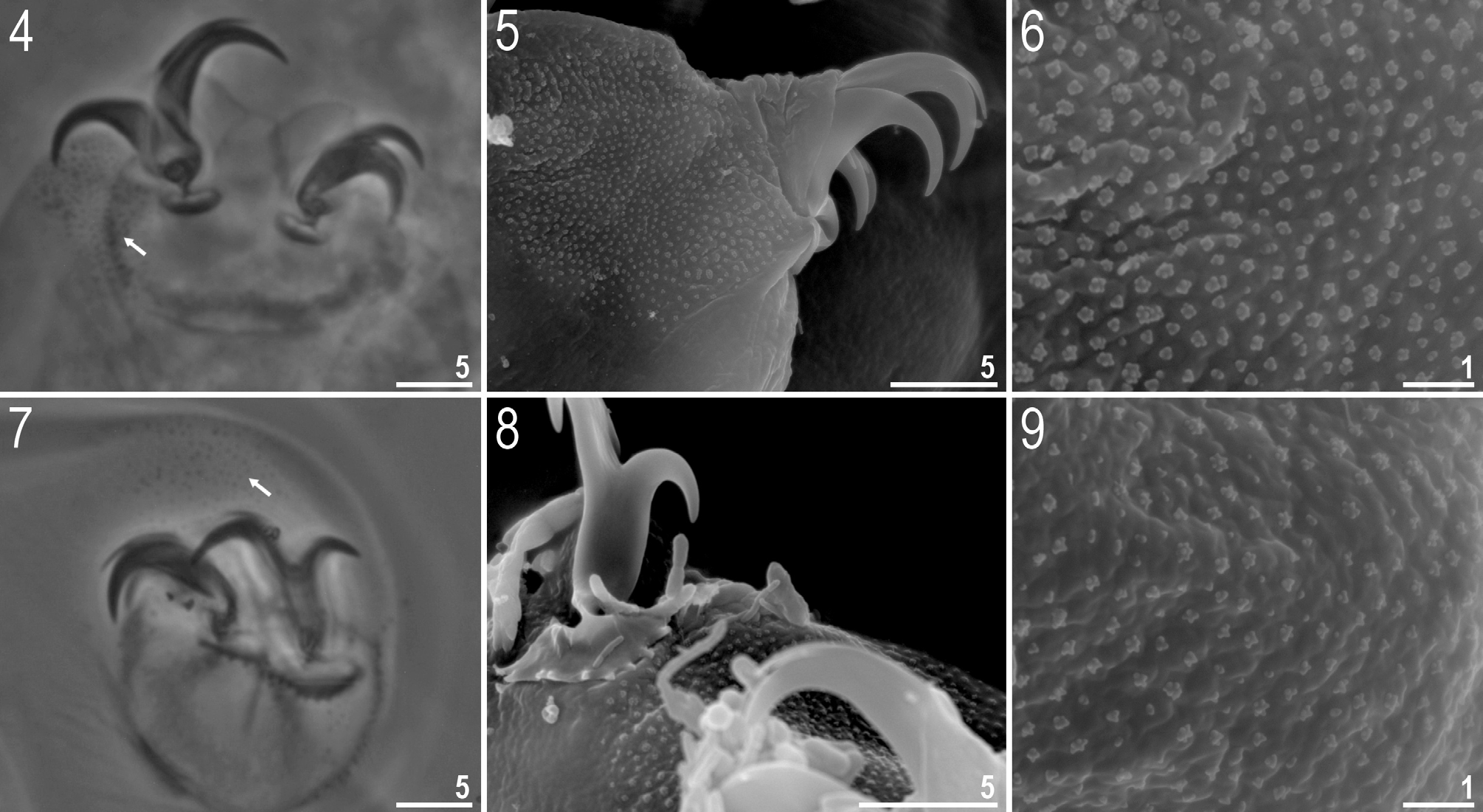
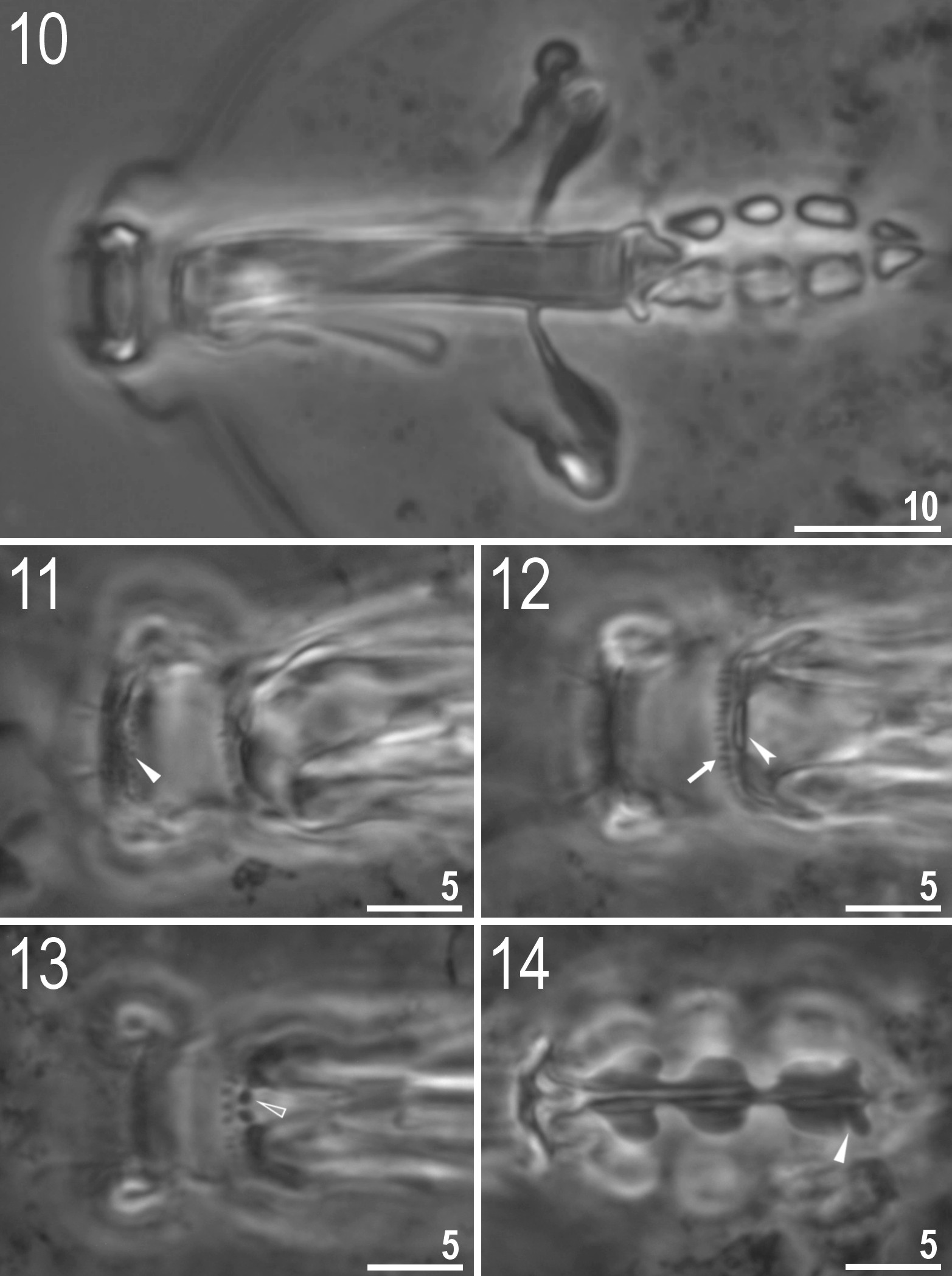
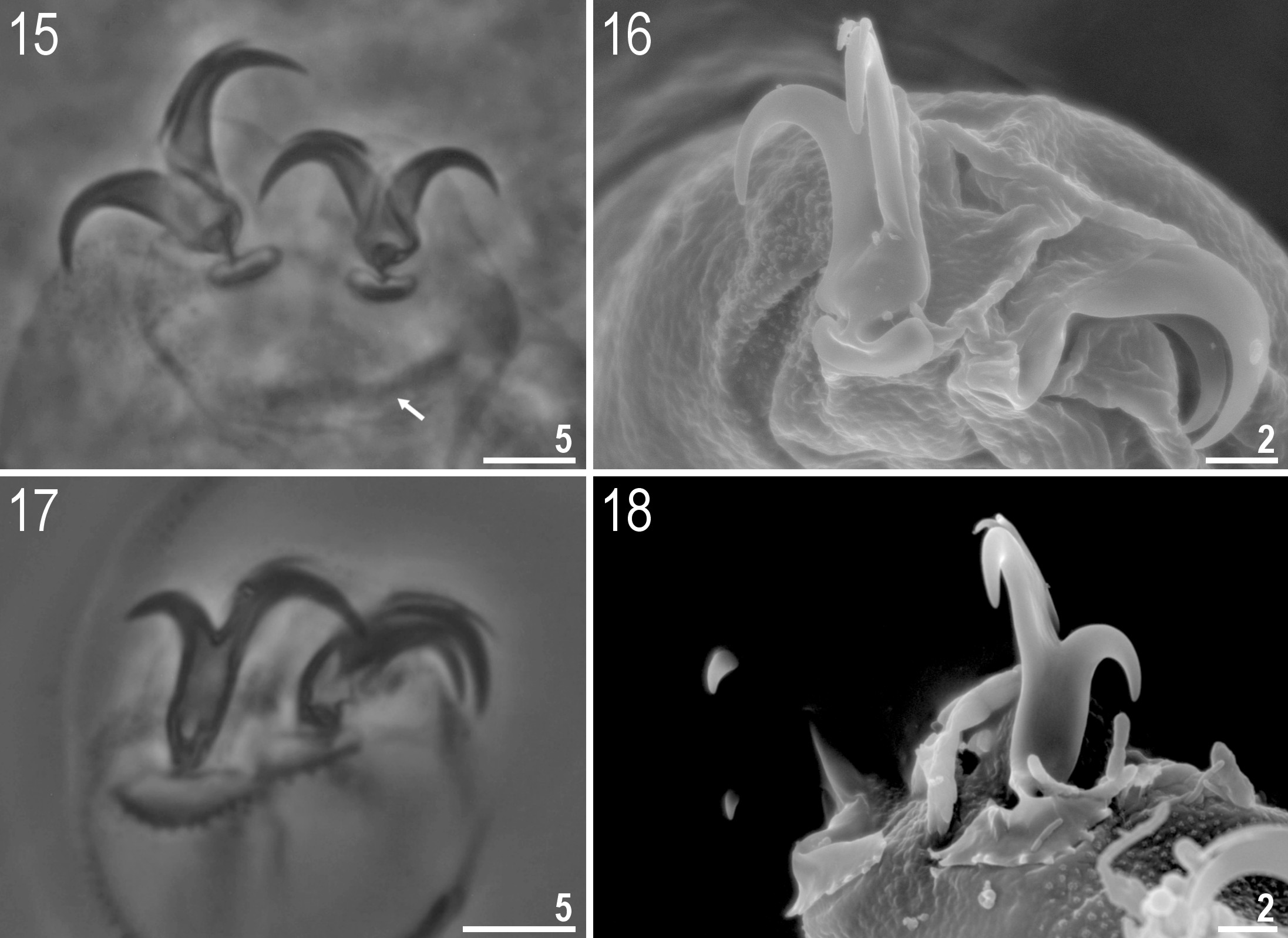
( Tables 3–4, Figs 1–32 View FIGURES 1–3 View FIGURES 4–9 View FIGURES 10–14 View FIGURES 15–18 View FIGURES 19–24 View FIGURES 25–32 )
Material examined: Holotype and 30 paratypes (19 animals and 11 eggs). Specimens mounted on microscope slides in Hoyer’s medium, fixed on SEM stubs or processed for DNA sequencing.
Description of the new species. Animals (measurements and statistics in Table 3). Body white in living specimens and transparent after fixation ( Figs 1–3 View FIGURES 1–3 ). Eyes absent after fixation in all studied specimens. Cuticle smooth, i.e. without gibbosities, papillae, spines, sculpturing or pores. Granulation present on external cuticle of all legs ( Figs 4–9 View FIGURES 4–9 ). Under SEM granulation visible as aggregates consisted of two to a dozen or so microgranules ( Figs 6, 9 View FIGURES 4–9 ).
Bucco-pharyngeal apparatus of the Macrobiotus type, with the ventral lamina and ten peribuccal lamellae ( Fig. 10 View FIGURES 10–14 ). Mouth antero-ventral. The oral cavity armature well developed and with three bands of teeth ( Figs 11–13 View FIGURES 10–14 ). The first band of teeth comprises numerous small granules arranged in a several rows situated anteriorly in the oral cavity, just behind the bases of the peribuccal lamellae ( Fig. 11 View FIGURES 10–14 , arrowhead). The second band of teeth, situated between the ring fold and the third band of teeth, contains ridges parallel to the main axis of the buccal tube that are larger than in the granules of the first band ( Fig. 12 View FIGURES 10–14 , arrow). The teeth of the third band are located within the posterior portion of the oral cavity, between the second band of teeth and the buccal tube opening ( Figs 12–13 View FIGURES 10–14 ). The third band of teeth are divided into the dorsal and the ventral portion. Under PCM, dorsal teeth are visible as two lateral and one median transverse ridges ( Fig. 12 View FIGURES 10–14 , indented arrowhead), whereas the ventral portion as two lateral ridges and two roundish median teeth ( Fig. 13 View FIGURES 10–14 , empty arrowhead). In addition, on the ventral side some barely visible teeth (in shape of granules in LM) are present between second and third rows of teeth. Pharyngeal bulb spherical, with triangular apophyses, three rod–shaped macroplacoids, and a triangular microplacoid. Macroplacoid length sequence 3±1>2. The first macroplacoid narrower anteriorly, the second without constrictions and the third with a sub-terminal constriction ( Fig. 14 View FIGURES 10–14 , arrowhead).
Claws of the hufelandi type ( Figs 15–18 View FIGURES 15–18 ); internal, external, anterior and posterior identical in shape. Primary branches with distinct accessory points. Lunules under claws I–III smooth and dentate under claws IV ( Figs 15– 18 View FIGURES 15–18 ). Thin cuticular bars under claws I–III present ( Fig. 15 View FIGURES 15–18 , arrow). Other cuticular structures on legs absent.
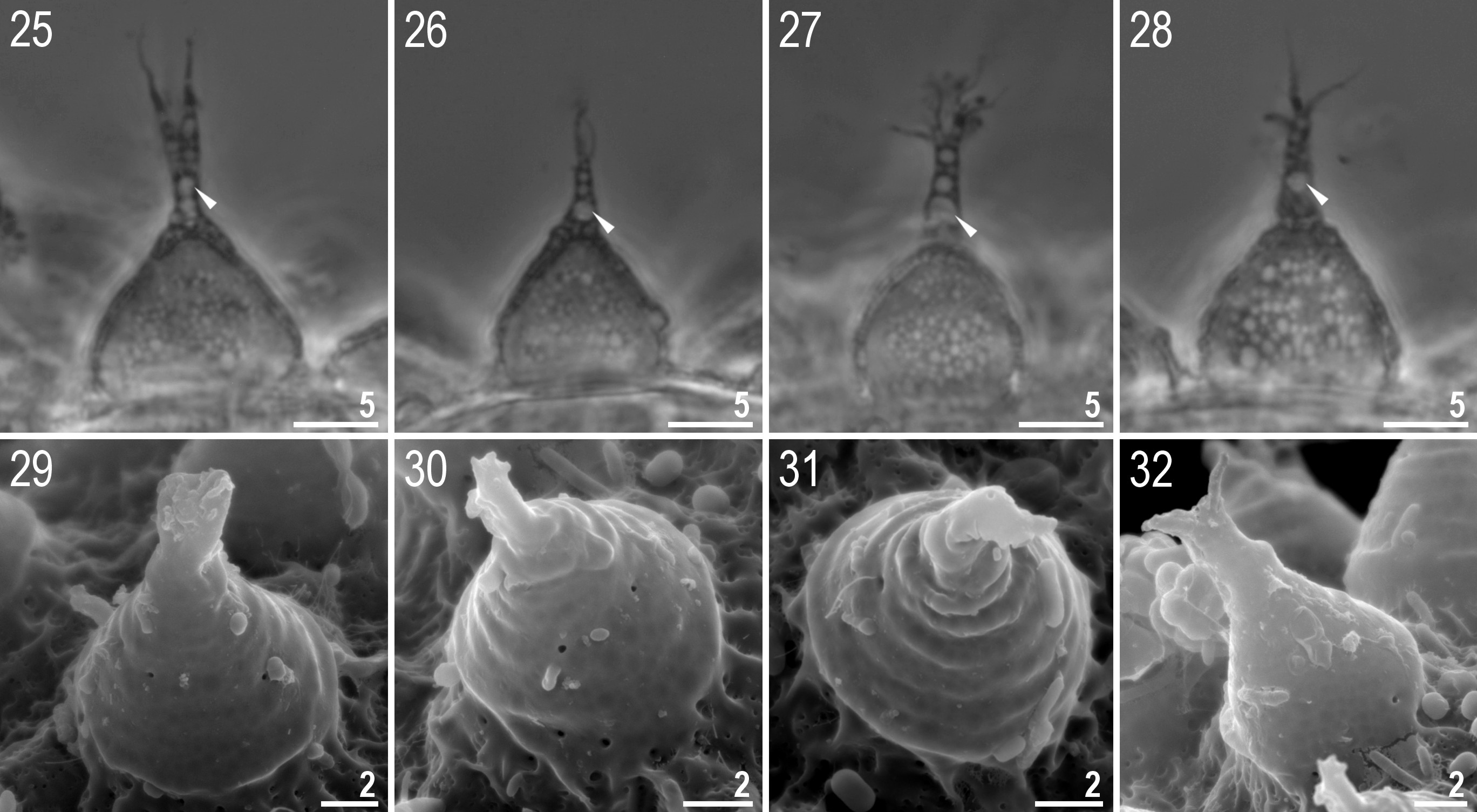
Eggs (measurements and statistics in Table 4). Laid freely, white, spherical and ornamented ( Figs 19–20 View FIGURES 19–24 ). Egg processes in the shape of wide, sharpened cones sometimes bifurcated at the tip or terminated by several short, thin and flexible filaments, visible both under PCM and SEM ( Figs 25–32 View FIGURES 25–32 ). The process surface is smooth with slight undulations poorly visible under PCM but clearer under SEM. Labyrinthine layer between process walls visible under PCM as a clear reticular pattern with the mesh (0.3–1.5 µm in diameter), slightly increased in size from the tip to the base ( Figs 25–28 View FIGURES 25–32 ). In some processes, a slightly larger, bubble-mesh are present near the tip ( Figs 25–28 View FIGURES 25–32 , arrowheads). Under SEM, clearly visible pores are present on the processes, but mainly toward the base ( Fig. 24 View FIGURES 19–24 , arrowheads). Each process surrounded by a crown of thickenings, which form small ribs and then wrinkles on the egg surface ( Figs 21, 22, 24 View FIGURES 19–24 , arrows). Under PCM, the surface between processes appears dotted or with small wrinkles ( Figs 21–22 View FIGURES 19–24 ), while under SEM, these are visible as small pores and ridges ( Fig. 24 View FIGURES 19–24 ).
DNA sequences. The DNA was successfully extracted from only one of the three individuals used for the analysis. Nevertheless, from this one paragenophore we obtained sequences of very good quality for all four of the previously mentioned molecular markers, which are as follow:
The 18S rRNA sequence (GenBank: MH197158 View Materials ), 1020 bp long;
The 28S rRNA sequence (GenBank: MH197151 View Materials ), 808 bp long;
The ITS-2 sequence (GenBank: MH197150 View Materials ), 486 bp long;
The COI sequence (GenBank: MH195149 View Materials ), 658 bp long;
Type locality: 00°45'34''N, 79°35'24W; 98 m asl: Tumbes-Chocó-Magdalena hotspot, Ecuador, Esmeraldas Province, next to E20 road to Quinindé, out of Chinca city, mixed moss and lichen on rock, collectors: Milena Roszkowska and Łukasz Kaczmarek.
Etymology: The first author takes great pleasure in dedicating this new species to her friend—Roman Tarasewicz.
Type depositories: Holotype: slide EC1319/6 (with six paratypes) and seven paratypes (two specimens and five eggs) (slides: EC1319/1, EC1319/2, EC1319/3, EC1319/8) are deposited at the Department of Animal Taxonomy and Ecology, Faculty of Biology, Institute of Environmental Biology , Adam Mickiewicz University, Poznań, Umultowska 89, 61-614 Poznań, Poland; nine paratypes (six specimens and three eggs) (slides: EC1319/5, EC1319/7, EC1319/9) are deposited at the Museo Ecuatoriano de Ciencias Naturales , Sección de Entomología , Rumipamba 341 y Av. de los Shyris , Quito , Ecuador .
Phenotypic differential diagnosis. Based on the presence of reticulated, conical egg processes surrounded by crown of thickening and the absence of areolation on the egg surface, M. romani sp. nov. is most similar to the following 10 species: M. binieki ( Kaczmarek, Gołdyn, Prokop & Michalczyk, 2011) , M. coronatus (de Barros, 1942) , M. dimentmani ( Pilato, Lisi & Binda, 2010) , M. patiens ( Pilato, Binda, Napolitano & Moncada, 2000) , M. perfidus ( Pilato & Lisi, 2009) , M. philippinicus Mapalo, Stec, Mirano-Bascos & Michalczyk, 2016 , M. pseudoblocki Roszkowska, Stec, Ciobanu & Kaczmarek, 2016 , M. pseudocoronatus ( Pilato, Binda & Lisi, 2006) , M. pseudopatiens Kaczmarek & Roszkowska, 2016 , M. radiatus ( Pilato, Binda & Catanzaro, 1991) , M. rigidus (Pilato & Lisi, 2006) , M. simulans ( Pilato, Binda, Napolitano & Moncada, 2000) and M. wuzhishanensis ( Yin, L. Wang & X. Li, 2011) . Despite the similarities, M. romani sp. nov differs specifically from:
M. binieki View in CoL , only reported from the type locality in Bulgaria ( Kaczmarek et al. 2011), by: the presence of dentate lunules IV, different macroplacoid length sequence (1>3> 2 in M. binieki View in CoL vs. 3±1> 2 in M. romani View in CoL sp. nov.), higher pt of: buccal tube external width, macroplacoid 2 length, microplacoid length, claw I–III primary and secondary external branches (see Tables 3 and 1 in Kaczmarek et al. (2011) for the exact differences), different shape of the egg processes (long, smooth flexible spines, with very wide bases in M. binieki View in CoL vs. typically developed cones in M. romani View in CoL sp. nov.), presence of bifurcated processes or with several short, thin, and flexible filaments at the tip, smaller egg diameter without processes (85.1–94.5 µm in M. binieki View in CoL vs. 62.0–85.0 µm in M. romani View in CoL sp. nov.), longer egg processes (9.8–14.5 µm in M. binieki View in CoL vs. 14.6–21.4 µm in M. romani View in CoL sp. nov.), wider egg process bases (6.5–9.0 µm in M. binieki View in CoL vs. 9.6–12.7 µm in M. romani View in CoL sp. nov.) and smaller number of processes on the egg circumference (27–32 in M. binieki View in CoL vs. 16–17 in M. romani View in CoL sp. nov.).
M. coronatus View in CoL , reported from a few localities in South America (de Barros 1942; Pilato et al. 2000; Kaczmarek et al. 2015), by: the absence of eyes*, presence of dentate lunules IV, different macroplacoid length sequence (1>3> 2 in M. coronatus View in CoL vs. 3±1> 2 in M. romani View in CoL sp. nov.), presence of several short, thin, and flexible filaments on the top of egg processes, larger egg diameter with and without processes (55.0–71.0 µm, and 42.0–55.0 µm respectively in M. coronatus View in CoL vs. 95.4–116.1 µm and 62.0–85.0 µm respectively in M. romani View in CoL sp. nov.), longer egg processes (ca. 9.2 µm in M. coronatus View in CoL vs. 14.6–21.4 µm in M. romani View in CoL sp. nov.).
M. dimentmani View in CoL , reported from the type locality in Israel (Pilato et al. 2010), by: teeth of the third band visible as two lateral ridges and two roundish median teeth (three ridges in M. dimentmani View in CoL ), first macroplacoid narrower anteriorly (central narrowing in M. dimentmani View in CoL ), anterior and posterior claws IV of a clearly different length, shorter anterior and posterior claws IV, and lower pt of anterior and posterior primary and secondary branches of claws IV (see Tables 3 and 4 in Pilato et al. (2010) for the exact differences).
M. patiens , recorded from a few localities in Italy ( Pilato et al. 2000), by: the presence of dentate lunules IV, and presence of bifurcated egg processes or with several short, thin, and flexible filaments at the tip.
M. perfidus , reported from three localities in the Seychelles ( Pilato & Lisi 2009), by: the presence of first row of teeth in oral cavity, the absence of tubercles on dorsal cuticle, absence of eyes*, presence of granulation on legs I–III, presence of dentate lunules IV, presence of bifurcated egg processes or with several short, thin, and flexible filaments at the tip, and higher number of processes on the egg circumference (11–13 in M. perfidus vs. 16–17 in M. romani sp. nov.).
M. philippinicus , reported from the type locality in Philippines ( Mapalo et al. 2016), by: the absence of eyes*, no granulation visible under SEM, different macroplacoid length sequence (1>3> 2 in M. philippinicus vs. 3±1> 2 in M. romani sp. nov.), absence of granulation on the filaments at the tip of the egg processes, and longer egg processes (2.1–13.7 µm in M. philippinicus vs. 14.6–21.4 µm in M. romani sp. nov.).
M. pseudoblocki , reported from the type locality in Argentina (Roszkowska et al. 2016), by: the absence of eyes*, presence of granulation on legs I–IV, different macroplacoid length sequence (1>3> 2 in M. pseudoblocki vs. 3±1> 2 in M. romani sp. nov.), presence of dentate lunules IV, larger internal secondary branch of claw III, higher pt of: stylet support insertion point, buccal tube external width, claw I–III external primary and internal secondary external branches, claw II–III external secondary branches (see Table 1 and 3 in Roszkowska et al. (2016) for the exact differences), larger egg diameter with processes (83.4–88.3 µm in M. pseudoblocki vs. 95.4–116.1 µm in M. romani sp. nov.), longer egg processes (10.5–12.8 µm in M. pseudoblocki vs. 14.6–21.4 µm in M. romani sp. nov.), wider egg process bases (5.8–7.6 µm in M. pseudoblocki vs. 9.6–12.7 µm in M. romani sp. nov.), and smaller number of processes on the egg circumference (20–24 in M. pseudoblocki vs. 16–17 in M. romani sp. nov.).
M. pseudocoronatus , reported from the type locality on Seychelles (Pilato et al. 2006), by: the absence of tubercles on dorsal cuticle, absence of eyes*, higher number of processes on egg circumference (ca. 14 in M. pseudocoronatus vs. 16–17 in M. romani sp. nov.), larger eggs with and without processes (ca. 82.3 and ca. 50.1 µm respectively in M. pseudocoronatus vs. 95.4–116.1 and 62.0–85.0 µm in M. romani sp. nov.), and longer egg processes (10.9–12.7 µm in M. pseudocoronatus vs. 14.6–21.4 µm in M. romani sp. nov).
M. pseudopatiens , reported from the type locality in Costa Rica (Kaczmarek & Roszkowska 2016), by: the presence of first row of teeth in oral cavity, different macroplacoid length sequence (1>3> 2 in M. pseudopatiens vs. 3±1> 2 in M. romani sp. nov.), presence of granulation on legs I–III, presence of dentate lunules IV, larger internal secondary branch of claw I and III, higher pt of: claw I internal secondary branches, claw II external primary and internal secondary branches, claw III external and internal secondary branches (see Table 1 and 3 in Kaczmarek & Roszkowska (2016) for the exact differences), higher number of processes on egg circumference (11–12 in M. pseudopatiens vs. 16–17 in M. romani sp. nov.), and larger egg diameter with and without processes (80.4–88.0 and 55.5–59.3 µm respectively in M. pseudopatiens vs. 95.4–116.1 and 62.0–85.0 µm in M. romani sp. nov.).
M. radiatus , reported only from Tanzania, Democratic Republic of Congo and Kenya ( Pilato et al. 1991, Stec et al. in review), by: smooth cuticle under SEM, absence of spurs on claws I–III, absence of pores on the top of egg processes, lack of microgranules on the filaments on the top of egg processes, higher number of processes on the egg circumference (10–12 in M. radiatus vs. 16–17 in M. romani sp. nov.), and narrower egg processes (14.5–22.5 in M. radiatus vs. 9.6–12.7 in M. romani sp. nov.).
M. rigidus , reported from the type locality in New Zealand (Pilato & Lisi 2006), by: the presence of granulation on legs I–III, the presence of dentate lunules IV, presence of bifurcated egg processes or with several short, thin, and flexible filaments at the tip, higher number of processes on egg circumference (ca. 12 in M. rigidus vs. 16–17 in M. romani sp. nov.), and larger egg diameter with processes (ca. 91.0 µm in M. rigidus vs. 95.4–116.1 µm in M. romani sp. nov.).
M. simulans , recorded from a few localities in Italy and Israel ( Pilato et al. 2000; Pilato et al. 2010), by: the absence of eyes*, presence of bifurcated processes or with several short, thin, and flexible filaments at the tip, longer egg processes (up to 11 µm in M. simulans vs. 14.6–21.4 µm in M. romani sp. nov.).
M. wuzhishanensis , reported from the type locality in China ( Yin et al. 2011), by: the absence of eyes*, different macroplacoid length sequence (3>1> 2 in M. wuzhishanensis vs. 3±1> 2 in M. romani sp. nov.), egg shell surface between processes dotted or with small wrinkles, and processes never trifurcated.
*character uncertain; Hoyer’s medium has the potential to cause eyes to ‘disappear’.
Genotypic differential diagnosis. The ranges of uncorrected genetic p-distances between the Mesobiotus romani sp. nov. and species of the genus Mesobiotus , for which sequences are available from GenBank (see Table 2 for details), are as follows (from the most to the least conservative):
18S rRNA: 1.1–5.9% (3.9% on average), with the most similar being M. philippinicus from Philippines ( KX129793 View Materials ) and the least similar being M. cf. mottai from Antarctica ( KT226072 View Materials );
28S rRNA: 6.8–9.9% (8.5% on average), with the most similar being M. philippinicus from Philippines ( KX129794 View Materials ) and the least similar being M. ethiopicus Stec & Kristensen, 2017 from Ethiopia ( MF678792 View Materials );
COI: 19.6–23.4% (21.7% on average), with the most similar being M. insanis Mapalo, Stec, Mirano-Bascos & Michalczyk, 2017 from Philippines ( MF441491 View Materials ) and the least similar being M. hilariae Vecchi, Cesari, Bertolani, Jönsoon, Rebecchi & Guidetti, 2016 from Antarctica ( KT226108 View Materials );
ITS-2: 25.5–28.6% (27.1% on average), with the most similar being M. philippinicus from Philippines ( KX129795 View Materials ) and the least similar being M. insanis from Philippines ( MF441490 View Materials ).

| DNA |
Department of Natural Resources, Environment, The Arts and Sport |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mesobiotus romani
| Roszkowska, Milena, Stec, Daniel, Gawlak, Magdalena & Kaczmarek, Łukasz 2018 |
M. romani
| Roszkowska & Stec & Gawlak & Kaczmarek 2018 |
M. romani
| Roszkowska & Stec & Gawlak & Kaczmarek 2018 |
M. romani
| Roszkowska & Stec & Gawlak & Kaczmarek 2018 |
M. romani
| Roszkowska & Stec & Gawlak & Kaczmarek 2018 |
M. romani
| Roszkowska & Stec & Gawlak & Kaczmarek 2018 |
M. romani
| Roszkowska & Stec & Gawlak & Kaczmarek 2018 |
M. romani
| Roszkowska & Stec & Gawlak & Kaczmarek 2018 |
M. romani
| Roszkowska & Stec & Gawlak & Kaczmarek 2018 |
M. romani
| Roszkowska & Stec & Gawlak & Kaczmarek 2018 |