
Argulus mongolianus Tokioka, 1939
|
publication ID |
https://doi.org/ 10.12782/specdiv.27.167 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA879F-FF87-405E-9CA5-FE395886F98E |
|
treatment provided by |
Felipe |
|
scientific name |
Argulus mongolianus Tokioka, 1939 |
| status |
|
Argulus mongolianus Tokioka, 1939
[Japanese name: Mouko-chou, based on Tokioka (1940)] ( Figs 1–6 View Fig View Fig View Fig View Fig View Fig View Fig )
Argulus mongolianus Tokioka, 1939: 42–47 , figs 1–2, pl. 1; Tokioka 1940: 306–308, figs 3–4; Yamaguti 1963: 325, pl. 323, fig. 2 (monograph); Song and Kuang 1980: 82, unnumbered figs (monograph); Kuang and Qian 1991: 166– 107, fig. 103 (monograph); Ky and Te 2007: 299, fig. 316; Wadeh et al. 2008: 765 (misspelled as A. mongolisnus ); Neethling and Avenant-Oldewage 2016: 1341 (monograph); Shedko et al. 2018: 70.
Hosts. Largemouth bass, Micropterus salmoides (Lacepède, 1802) (Centrarchiformes: Centrarchidae ); common carp, Cyprinus carpio Linnaeus, 1758 ( Cypriniformes : Cyprinidae ); Japanese white crucian carp, Carassius cuvieri Temminck and Schlegel, 1846 ( Cypriniformes : Cyprinidae ); and silver crucian carp, Carassius sp. ( Cypriniformes : Cyprinidae ).
Attachment sites. Buccal cavity wall, body surface.
Localities. Lake Izunuma (38º43′09″N, 141º06′09″E at center) and Lake Uchinuma (38º42′41″N, 141º04′33″E at center), Miyagi Prefecture, Japan GoogleMaps .
Material examined. One adult female, NSMT-Cr 29369, and three adult males, NSMT-Cr 29370, from the buccal cavity wall of M . salmoides from Lake Izunuma on 2 June 2020; five adult females, NSMT-Cr 29371, and five adult males, NSMT-Cr 29372, from the buccal cavity wall of M . salmoides from Lake Izunuma on 12 May 2020; two adult females and two adult males, NSMT-Cr 29373, from the buccal cavity wall of Cy . carpio from Lake Uchinuma on 12 May 2020; two adult females and one adult male, NSMTCr 29374, from the body surface of Ca . cuvieri from Lake Uchinuma on 12 May 2020; one adult male, NSMT-Cr 29375, from the body surface of Carassius sp . from Lake Uchinuma on 28 April 2020 .
Redescription. Based on six adult female specimens from M. salmoides . Body dorsoventrally flattened, measuring 6.6–8.3 (mean=7.3, n=6) mm in total length (from anterior tip of carapace to posterior tip of abdomen) and 4.0–4.7 (mean=4.4, n=6) mm in maximum width (around midlength of carapace).
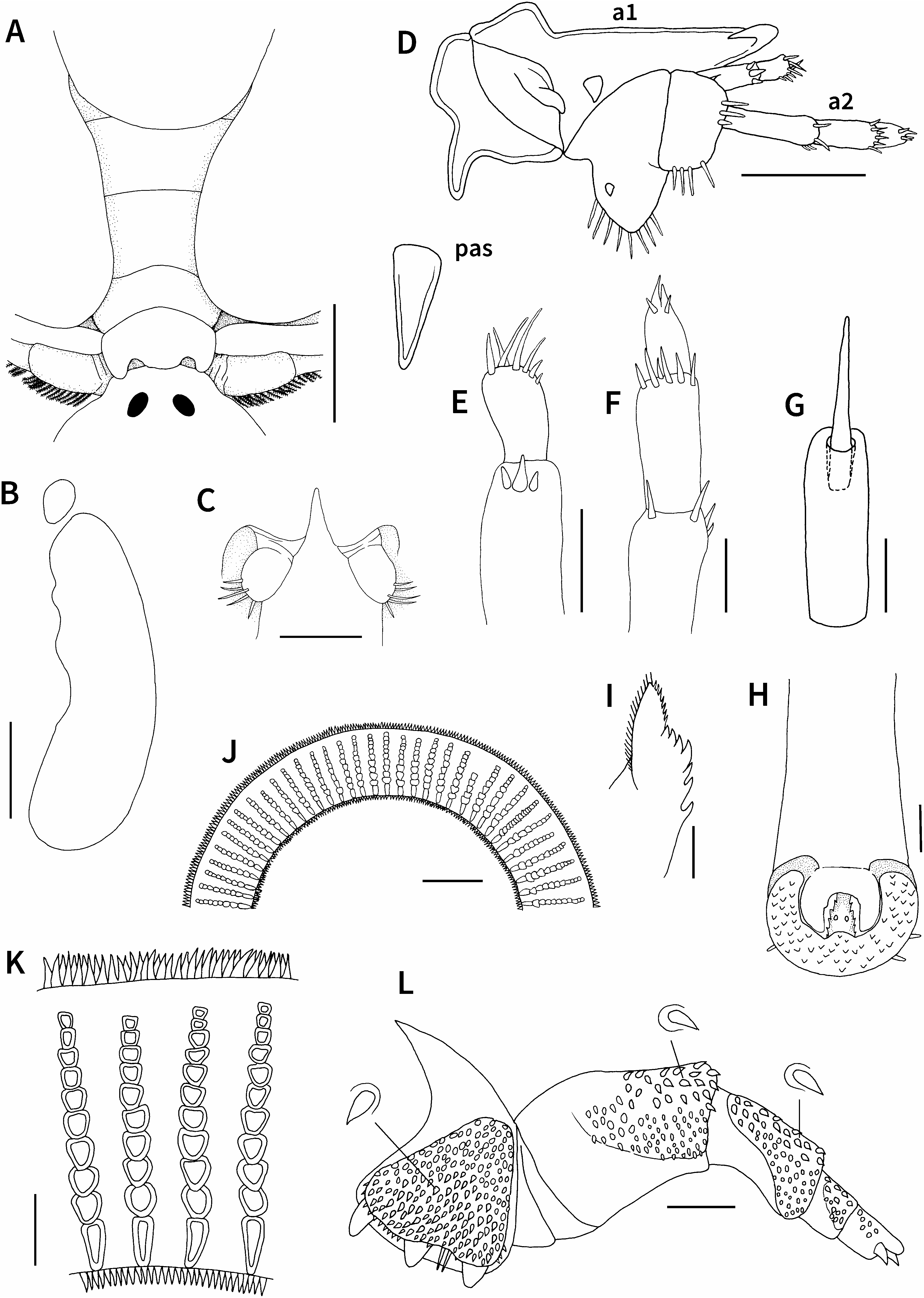
Carapace (including posterolateral lobes) circular, covering sympods and proximal part of first to third (or fourth in large specimens) pairs of legs in dorsal view, 4.3–5.5 (mean= 4.7, n=6) mm long, comprising 62.5–66.3% (mean=64.8%, n=6) of total length ( Figs 1A, B View Fig , 6A, B View Fig ). Centrofrontal region of carapace protruding anteriorly; anterior margin horizontal. Anterolateral indentations shallow; central longitudinal ribs distinct, anteriorly bifurcated. Compound eyes distinct, located dorsally at level where frontal region joins main part of carapace. Nauplius eye located posterior to compound eyes along midline of carapace. Dorsal surface of carapace smooth without spines. Ventral surface of lateral region of carapace ornamented with numerous, small posteriorly directed spines ( Fig. 1B View Fig ). Posterolateral lobes of carapace partially overlapped ( Fig. 1B View Fig ) or not overlapped ( Fig. 2A View Fig ), ending in rounded margin, separated by sinus nearly 40% length of carapace. Respiratory areas comprising small, oval anterior area and large, reniform posterior area; former area located at level between first and second maxillae, and latter at level of second maxillae to third pair of legs ( Figs 1B View Fig , 2B View Fig ). Thorax with four segments, bearing small spiniform projections on ventral surface; third segment narrower than first and second segments ( Fig. 1A, B View Fig ); fourth segment narrowest, with two posteriorly directed lateral protrusions and weakly convex posterior margin in dorsal view ( Fig. 2A View Fig ). Abdomen longer than wide; anal indentation 60.0–77.3% (mean=70.5%, n=6) of abdomen length forming two lobes; lateral margins and posterior surface of lobes with small sharply pointed spines; each lobe oval, becoming wider toward midlength, then narrower posteriorly, ending in rounded margin ( Fig. 1A, B View Fig ). Paired spermathecae each oval, located anteriorly in abdomen ( Figs 1A, B View Fig , 2A View Fig ). Caudal rami located at base of anal indentation, each with five naked setae on posterior margin ( Figs 1A View Fig , 2C View Fig ).
First antennae with four segments ( Fig. 2D, E View Fig ): first segment heavily sclerotized in mesial and posterior regions, with large projection on mesial margin; second segment also heavily sclerotized, with triangular projection on anterior margin, strong apically bent hook at distal corner, stout projection near posterior margin, and oblique swelling near mesial margin; third segment cylindrical, with three naked setae, one of them stout and larger than others; apical segment shorter than third segment, with seven naked setae of different sizes at tip. Second antennae with five segments ( Fig. 2D, F View Fig ): first segment sclerotized, with large rounded protrusion bearing eight naked setae on posterior margin and short spine in centroventral region; second segment shorter than first segment, with four naked setae on posterior margin, three naked setae near distal margin, and short seta on anterior margin; third, fourth, and apical segments nearly cylindrical and decreasing in length, possessing four, six, and four naked setae, respectively. Postantennal spines large and robust, each located posterior to large projection of first segment of first antenna ( Figs 1B View Fig , 2D View Fig ). Preoral sheath cylindrical and visible near anterior regions of first maxillae on ventral midline of carapace ( Figs 1B View Fig , 2G View Fig ); anterior portion of stylet protruding from opening of preoral sheath. Mouth tube located just posterior to preoral sheath, longer than wide, becoming gradually wider posteriorly ( Figs 1B View Fig , 2H View Fig ); mouth composed of small anterior serrated labrum and larger posterior labium with scales on ventral surface and several short naked setae on outer margin ( Fig. 2H View Fig ); mandibles completely covered by labrum, composed of anterior part bearing many tiny spines on both distal and mesial sides and posterior part with seven teeth on mesial side ( Fig. 2I View Fig ); small paired buccal spines present on ventral surface of labium ( Fig. 2H View Fig ).
First maxillae forming well developed cup-like suckers ( Fig. 1B View Fig ), with 64–71 (mean=68, n=10) supporting rods in sucker membrane ( Fig. 2J View Fig ). Supporting rods each composed of one basal elongate plus eight to ten trapezoidal sclerites, slightly wider at second to fourth sclerites, and tapering distally [of 35 supporting rods examined, commonest number of sclerites per rod is 10 (n=15), followed by 11 and nine (n=10 and 8, respectively), and two rods have unusual numbers of sclerites (one has six, the other 16 sclerites)] ( Fig. 2K View Fig ). Both outer and inner margins of rim of sucker membrane with numerous apically pointed projections ( Fig. 2J, K View Fig ). Second maxillae with five segments ( Fig. 2L View Fig ); first segment robust, with three basally separated, almost equally long blunt projections; corpus of first segment with two posteriorly directed naked spiniform setae between two projections and furnished with raised patch of numerous posteriorly directed scales; second segment longer than first segment, bearing some distally directed scales on anteroventral surface and small posteriorly directed scales on ventral surface; third segment shorter and narrower than second segment, with raised patch of distally directed scales on ventral surface (scales near anterior margin are slightly larger than those in other regions); fourth segment subquadrate, with raised patch of distally directed scales on ventral surface; terminal segment bearing some tiny scales on ventral surface, ending in one club-shaped protrusion and two hooklike claws. Accessory spines near ventral midline, slightly apart from first segments of second maxillae ( Fig. 1B View Fig ). Postmaxillary spines small and located just anterior to first segment of thorax ( Fig. 1B View Fig ).
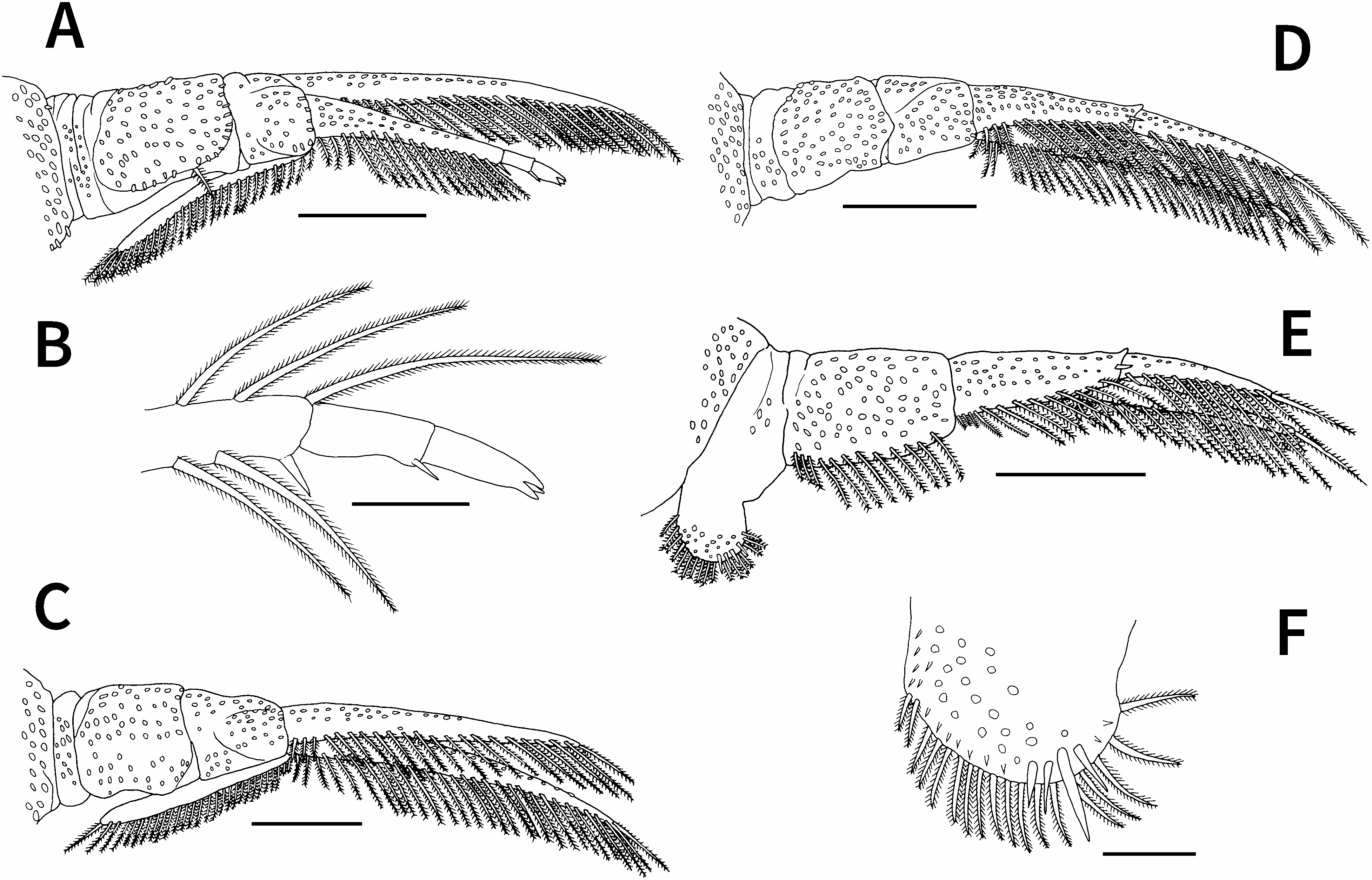
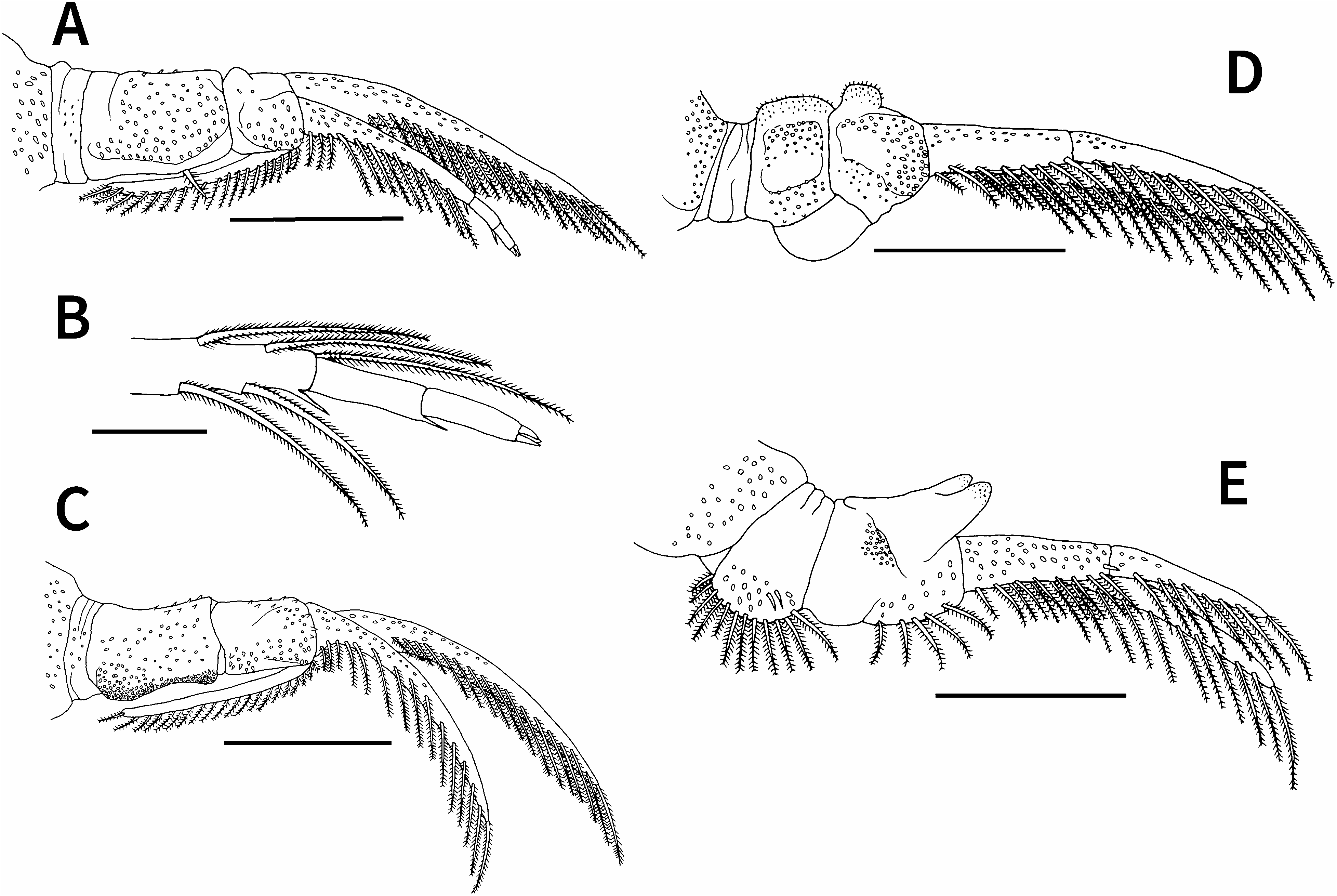
First to fourth pairs of legs ( Fig. 3 View Fig ) biramous, each with sympods composed of coxa and basis, sympods and rami of first to fourth legs ventrally covered with small spiniform projections; rami bearing two lateral rows of plumose setae each near ventro- and dorsoposterior margins; first and second pairs of legs each possessing dorsal flagellum projecting from extreme proximal part of exopod. First leg ( Fig. 3A, B View Fig ) coxa bearing single plumose seta near posterior margin; basis nearly half as long as coxa, with small swelling near anterior junction of coxa and basis; exopod unsegmented, with 23 plumose setae near ventroposterior margin; endopod three-segmented, proximal segment with 16 plumose setae near ventroposterior margin and short naked seta at posterodistal corner, middle segment with short naked seta at posterodistal corner, and terminal segment ending in three short spines; flagellum extending to proximal margin of coxa, with 28 plumose setae on posterior margin and no plumose setae on anterior margin. Second leg ( Fig. 3C View Fig ) sympod without setae on posterior margin; posterior margin of coxa weakly concave; basis slightly shorter than coxa; exopod and endopod unsegmented, with 30 and 22 plumose setae, respectively, near ventroposterior margin; flagellum extending to proximal margin of coxa, with 28 plumose setae on posterior margin and no plumose setae on anterior margin. Third leg ( Fig. 3D View Fig ) sympod without setae on posterior margin; basis slightly shorter than coxa; exopod unsegmented, with 29 plumose setae near ventroposterior margin; endopod two-segmented, proximal segment with 13 plumose setae near ventroposterior margin and short naked seta near distal margin, terminal segment with 11 plumose setae near ventroposterior margin. Fourth leg ( Fig. 3E, F View Fig ) coxa forming natatory lobe bearing 17 plumose setae on posterior margin and four short naked setae near posterior margin; basis longer than coxa, with 12 plumose setae near ventroposterior margin; exopod unsegmented, with 27 plumose setae near ventroposterior margin; endopod twosegmented, proximal segment with 13 plumose setae near ventroposterior margin and short naked seta near distal margin, terminal segment with 11 plumose setae and short naked seta near ventroposterior proximal margin.
Description of adult male. Based on eight specimens from M. salmoides . Body dorsoventrally flattened, measuring 4.2–5.6 (mean=4.8, n=8) mm in total length and 2.5– 3.2 (mean=2.8, n=8) mm in maximum width.
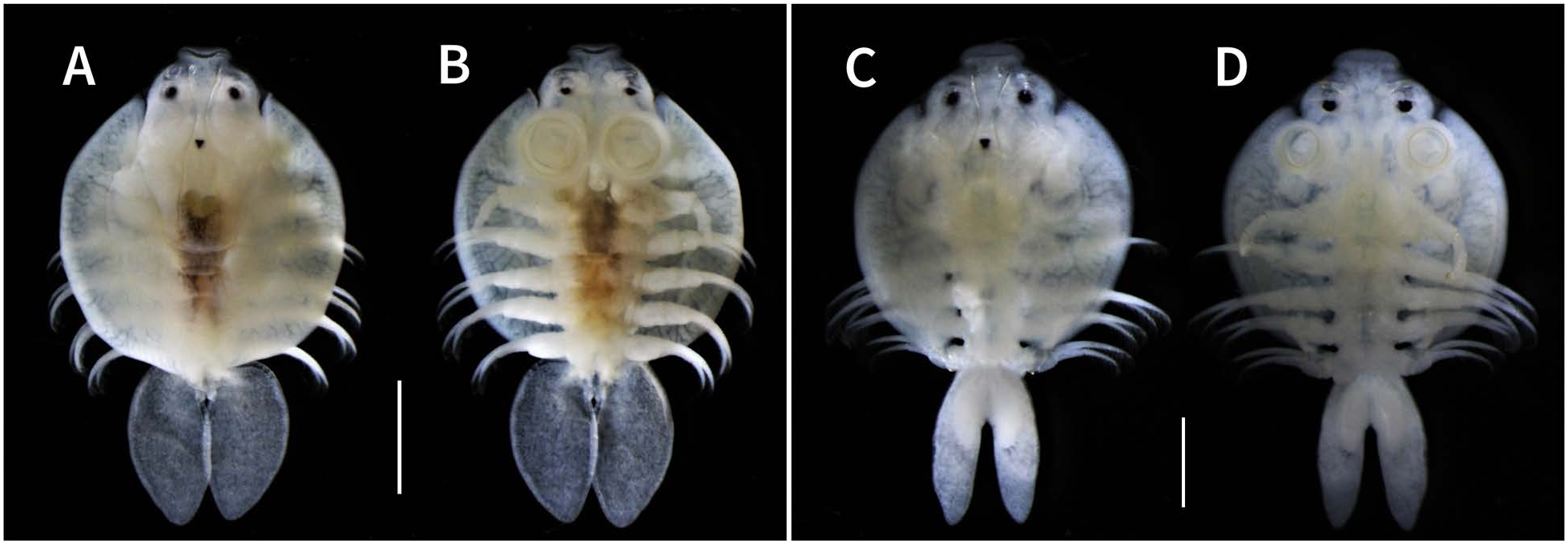
Carapace (including posterolateral lobes) nearly circular, covering sympods and proximal portions of first to third pairs of legs in dorsal view, 2.8–3.9 (mean=3.2, n=8) mm long, comprising 63.0–69.6% (mean=66.1%, n=8) of total length ( Figs 4A, B View Fig , 6C, D View Fig ). Centrofrontal region of carapace protruding anteriorly; anterior margin widely rounded or horizontal. Anterolateral indentations shallow; central longitudinal ribs distinct, anteriorly bifurcated. Compound eyes distinct, dorsally located at level where frontal region joins main portion of carapace. Nauplius eye located posterior to compound eyes along midline of carapace. Dorsal surface of carapace smooth without spines. Ventral surface of lateral region of carapace ornamented with numerous, small posteriorly directed spines ( Fig. 4B View Fig ). Posterolateral lobes of carapace not overlapped, ending in rounded margin, separated by sinus nearly 1/3 length of carapace. Respiratory areas comprising small, oval anterior area and large, reniform posterior area; former area located at level between first and second maxillae, and latter at level of second maxillae to third pair of legs ( Fig. 4B, C View Fig ). Thorax with four segments, bearing small spiniform projections on ventral surface; third segment shorter and narrower than first and second segments; fourth segment shortest and narrowest, ending in weakly concave posterior margin ( Fig. 4A, B View Fig ). Abdomen longer than wide; anal indentation 42.9–61.1% (mean=51.8%, n=8) of abdomen length forming two lobes; lateral margins and posterior surface of lobes with small sharply pointed spines; each lobe oblong, ending in rounded margin ( Fig. 4A, B View Fig ). Testes elliptical, located in anterior to middle portion of abdomen ( Fig. 4A, B View Fig ). Caudal rami locat- ed at base of anal indentation, each with five naked setae on posterior margin ( Fig. 4D View Fig ).
First antennae with four segments ( Fig. 4E, F View Fig ): first segment heavily sclerotized in mesial and posterior regions, with large projection on mesial margin; second segment also heavily sclerotized, with triangular projection on anterior margin, strong apically bent hook at distal corner, stout projection near posterior margin, and oblique swelling near mesial margin; third segment cylindrical, with three naked setae, one of them stout and larger than others; apical segment shorter than third segment, with seven naked setae of different sizes at tip. Second antennae with five segments ( Fig. 4E, G View Fig ): first segment sclerotized and ovoid, with large rounded protrusion bearing eight naked setae on posterior margin and short spine in centroventral region; second segment shorter than first segment, with four naked setae on posterior margin, three naked setae near distal margin, and short seta on anterior margin; third, fourth, and apical segments nearly cylindrical and decreasing in length, possessing four, six, and four naked setae, respectively. Postantennal spines large and robust, each located posterior to large projection of first segment of first antenna ( Fig. 4E View Fig ). Preoral sheath cylindrical, located between first maxillae on ventral midline of carapace ( Fig. 4B, H View Fig ); anterior portion of stylet protruding from opening of preoral sheath. Mouth tube located just posterior to preoral sheath, longer than wide, becoming gradually wider posteriorly ( Fig. 4B, I View Fig ); mouth composed of small anterior serrated labrum and larger posterior labium with scales on ventral surface and several short naked setae on outer margin ( Fig. 4I View Fig ); mandibles completely covered by labrum, composed of anterior part bearing many tiny spines on both distal and mesial sides and posterior part with five teeth on mesial side ( Fig. 4J View Fig ); small buccal paired spines present on ventral surface of labium ( Fig. 4I View Fig ).
First maxillae forming cup-like suckers ( Fig. 4B View Fig ), with 58–64 (mean=61, n=10) supporting rods in sucker membrane. Supporting rods each composed of one basal elongate plus six to eight trapezoidal sclerites, slightly wider at second to fourth sclerites, and tapering distally [of 30 supporting rods examined, commonest number of sclerites per rod is eight (n=17), followed by seven and nine (n=8 and 5, respectively)] ( Fig. 4K View Fig ). Both outer and inner margins of rim of sucker membrane with numerous apically pointed projections ( Fig. 4K View Fig ). Second maxillae with five segments ( Fig. 4L View Fig ); first segment robust, with three basally separated, almost equally long blunt projections; corpus of first segment with two posteriorly directed naked spiniform setae between two projections and furnished with raised patch of numerous posteriorly directed denticles; second segment longer than first segment, bearing some apically bifurcated denticles on anteroventral surface, some posteriorly directed denticles on centroventral surface, and small denticles on posteroventral surface; third segment shorter and narrower than second segment, with raised patch of some apically bifurcated denticles on anterodorsal surface and small distally or posteriorly directed denticles on other ventral surface; fourth segment subquadrate, with raised patch of distally directed denticles on ventral surface; terminal segment bearing some tiny denticles on ventral surface, ending in one club-shaped and two hook-like projections. Accessory spines near ventral midline, slightly apart from first segments of second maxillae ( Fig. 4B View Fig ). Postmaxillary spines small and located anterior to first segment of thorax ( Fig. 4B View Fig ).
First to fourth pairs of legs ( Fig. 5 View Fig ) biramous, with each sympod composed of coxa and basis; sympods and rami of first to fourth legs ventrally covered with small spiniform projections; rami bearing two lateral rows of plumose setae each near ventro- and dorsoposterior margins; first and second pairs of legs each possessing dorsal flagellum projecting from extreme proximal part of exopod. First leg ( Fig. 5A, B View Fig ) coxa bearing single plumose seta near posterior margin; basis nearly half as long as coxa, with small swelling near anterior junction of coxa and basis; exopod unsegmented, with 20 plumose setae near ventroposterior margin; endopod three-segmented, proximal segment with 12 plumose setae near ventroposterior margin and short naked seta at posterodistal corner, middle segment with short naked seta at posterodistal corner, terminal segment ending in three short spines; flagellum extending to proximal margin of coxa, with 18 plumose setae on posterior margin and no plumose setae on anterior margin. Second leg ( Fig. 5C View Fig ) sympod without setae on posterior margin; posterior margin of coxa weakly concave, densely armed with small projections; basis slightly shorter than coxa; exopod and endopod unsegmented, with 17 and 15 plumose setae, respectively, near ventroposterior margin; flagellum extending to proximal margin of coxa, with 20 plumose setae on posterior margin and no plumose setae on anterior margin. Third leg ( Fig. 5D View Fig ) sympod without setae on posterior margin, possessing swelling (=socket) at posterior junction of coxa and basis; basis nearly as long as coxa, bearing apically rounded projection on anterior margin; exopod unsegmented, with 18 plumose setae near ventroposterior margin; endopod twosegmented, proximal and terminal segments with eight and nine plumose setae, respectively, near ventroposterior margin. Fourth leg ( Fig. 5E View Fig ) coxa forming natatory lobe bearing 12 plumose setae on posterior margin and two short naked setae on ventroposterior surface; basis longer than coxa, bearing stout peg with distally bifurcated, rounded ends covered by minute scales on anterior margin and five plumose setae on posterior margin; exopod unsegmented, with 15 plumose setae near ventroposterior margin; endopod two-segmented, proximal segment with eight plumose setae near ventroposterior margin and short naked seta near distal margin, terminal segment with eight plumose setae near ventroposterior margin.
Color (based on ethanol-preserved specimens). Carapace, legs, and male thorax white; female thorax and centroposterior region of cephalothorax pale or dark yellow with irregularly shaped black spots unevenly scattered on dorsal surface; respiratory areas fringed weakly by black pigment ( Fig. 6 View Fig ).
Remarks. Argulus mongolianus was originally described based on a single ovigerous female from Lake Dalai-nor, Inner Mongolia, China ( Tokioka 1939). The host was not reported. Subsequently, A. mongolianus was reported from two provinces (Bắc Kạn and Lào Cai), northern Vietnam ( Ky and Te 2007), Guangdong Province, China ( Wadeh et al. 2008), and the Primorye Territory, Russian Far East ( Shedko et al. 2018) but no further description has been made to date. Tokioka (1940) published a report in which the original description of the species was translated from English into Japanese.
The adult females of the argulid collected in the present study are almost completely identical to the adult female of A. mongolianus described by Tokioka (1939). The adult males were also collected and as described above, their morphology is quite similar to that of the females. Therefore, both the females and males collected in this study are identified as A. mongolianus . The male is described for the first time. The species is characterized by the anterior protrusion of the centrofrontal carapace in both sexes ( Figs 1A, B View Fig , 4A, B View Fig , 6A–D View Fig ), a circular carapace and oval abdominal lobes in the female ( Figs 1A, B View Fig , 6A, B View Fig ), oblong abdominal lobes in the male ( Figs 4A, B View Fig , 6C, D View Fig ), and the presence of a single plumose seta near the posterior margin of the coxa of the first pair of legs in both sexes ( Figs 3A View Fig , 5A View Fig ).
There are several differences in the morphology of A. mongolianus between the original and present descriptions. The ventral surface of the frontal region of the carapace of the female was figured to have many small spines ( Tokioka 1939: pl. 3, fig. A), but both the female and male of the species examined in this study have no such spines. While the dorsoposterior margins of the female’s thoracic segments were reported to be “complexly corrugated” ( Tokioka 1939: pl. 3, fig. C), no similar structure was observed in the specimens of both sexes examined in this study. Further, the number of supporting rods in the sucker membrane of the female was reported to be “70–80” ( Tokioka 1939), but the females and males examined herein have fewer [64–71 (mean=68) and 58–64 (mean=61)] supporting rods, respectively. It seems likely that Tokioka (1939) incorrectly counted the number of supporting rods.
There are morphological differences between the sexes of A. mongolianus . The female’s carapace is more circular than the male’s, and the female’s abdominal lobe is oval but the male’s is oblong ( Figs 1A, B View Fig , 4A, B View Fig , 6A–D View Fig ). Only the male has accessory copulatory structures, i.e., the socket and peg on the third and fourth pairs of legs, respectively ( Fig. 5D, E View Fig ). The male possesses both posteriorly directed and apically bifurcated scales on the second and third segments of the second maxilla ( Fig. 4K View Fig ), but the female has only posteriorly directed scales on these segments ( Fig. 2K View Fig ). The posterior margin of the coxa of the male’s second leg is densely armed with small projections ( Fig. 5C View Fig ), but the female has no such ornamentation on the same margin ( Fig. 3C View Fig ). Moreover, as stated above, the female’s first maxilla has more supporting rods (64–71) than the male’s (58–64). Similarly, the number of sclerites per supporting rod in the female (9–11) is more numerous than in the male (7–9). These sex-related differences in the number of supporting rods and sclerites per rod are probably related to differences in the carapace length of the females [4.3–4.8 (mean=4.6, n=5) mm] and males [2.8–3.2 (mean=2.9, n=5) mm] examined. It is known, for example, that larger individuals of Argulus melanostictus C. B. Wilson, 1935 have more supporting rods and sclerites per rod than smaller ones ( Benz et al. 1995).
As reported by Boxshall and Jaume (2009) in A. japonicus , A. mongolianus also has a flagellum projecting from the extreme proximal part of the exopod of each of the first and second pairs of legs ( Figs 3A, B View Fig , 5A, B View Fig ).
Information on the hosts of A. mongolianus is quite limited. In the original description of the species from Lake Dai-nor, Inner Mongolia, the host was reported as unknown ( Tokioka 1939). Wadeh et al. (2008) later listed A. mongolianus as one of eight congeneric species infecting farmed fishes in Guangdong Province, China, but no fish name was reported. In contrast, two and one species of fishes have been reported as the hosts of A. mongolianus , respectively, from northern Vietnam ( Ky and Te 2007) and Russian Far East ( Shedko et al. 2018): the two hosts are Spinibarbus denticularis (Oshima, 1926) ( Cypriniformes : Cyprinidae ) and Bagarius bagarius (Hamilton, 1822) ( Siluriformes : Sisoridae ), and another host is common carp. In the present study, A. mongolianus was collected from largemouth bass, common carp, Japanese white crucian carp, and silver crucian carp. Excluding common carp, the three other species represent new host records. Based on the host species reported in this and previous papers ( Ky and Te 2007; Shedko et al. 2018), A. mongolianus is not strictly host-specific but may commonly utilize cyprinids as its hosts.
Largemouth bass was introduced in 1925 and 1972 from North America into Japan ( Takamura 2007), where this fish species is known as a host of A. japonicus as well (Nagasawa 2021). Similarly, A. japonicus has been reported in Japan from common carp ( Nagasawa et al. 2012, 2013; Nagasawa 2018; Nagasawa and Ishiyama 2019; see Nagasawa 2009, 2011a for the earlier literature), Japanese white crucian carp ( Takeda et al. 2000; Nagasawa 2011b; Nagasawa et al. 2012), and silver crucian carp ( Takeda et al. 2000, reported as Ca. auratus langsdorfii ; Nagasawa 2011b, reported as Ca. a. langsdorfii).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Argulus mongolianus Tokioka, 1939
| Nagasawa, Kazuya, Asayama, Takato & Fujimoto, Yasufumi 2022 |
Argulus mongolianus
| Shedko, M. B. & Ermolenko, A. V. & Aseeva, N. L. 2018: 70 |
| Neethling, L. A. & Avenant-Oldewage, A. 2016: 1341 |
| Wadeh, H. & Yang, J. W. & Li, G. Q. 2008: 765 |
| Ky, H. & Te, B. Q. 2007: 299 |
| Kuang, P. & Qian, J. 1991: 166 |
| Song, D. S. & Kuang, P. R. 1980: 82 |
| Yamaguti, S. 1963: 325 |
| Tokioka, T. 1940: 306 |
| Tokioka, T. 1939: 47 |