
Astyanax angustifrons ( Regan 1908 )
|
publication ID |
https://doi.org/ 10.1080/00222933.2017.1324050 |
|
publication LSID |
lsid:zoobank.org:pub:ABC57223-DF66-49B6-8FE0-87CFF5D3EA03 |
|
persistent identifier |
https://treatment.plazi.org/id/03806F39-C94B-FFF8-FE0A-FA76D2B4FB02 |
|
treatment provided by |
Felipe |
|
scientific name |
Astyanax angustifrons ( Regan 1908 ) |
| status |
|
Astyanax angustifrons ( Regan 1908) View in CoL
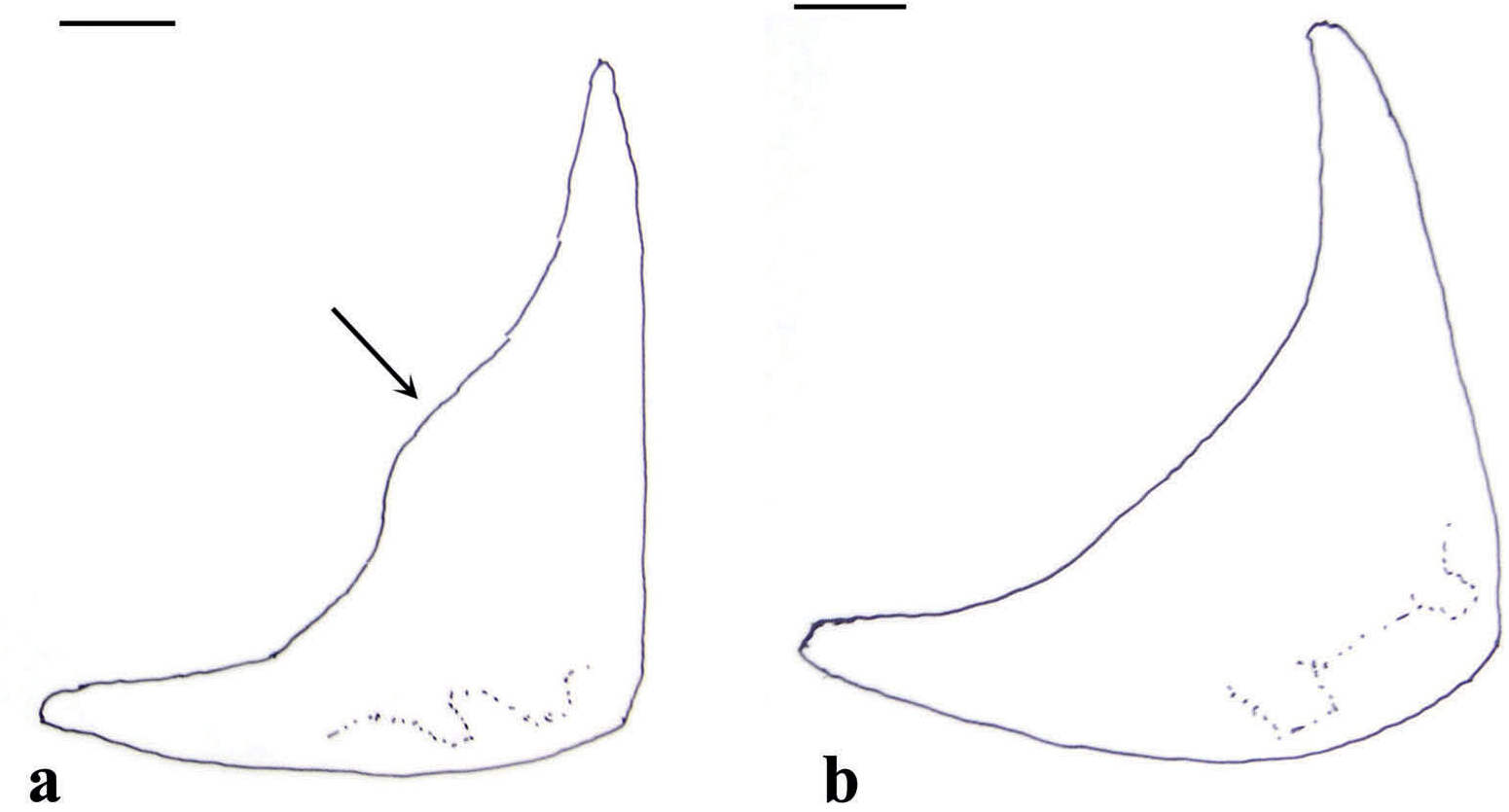
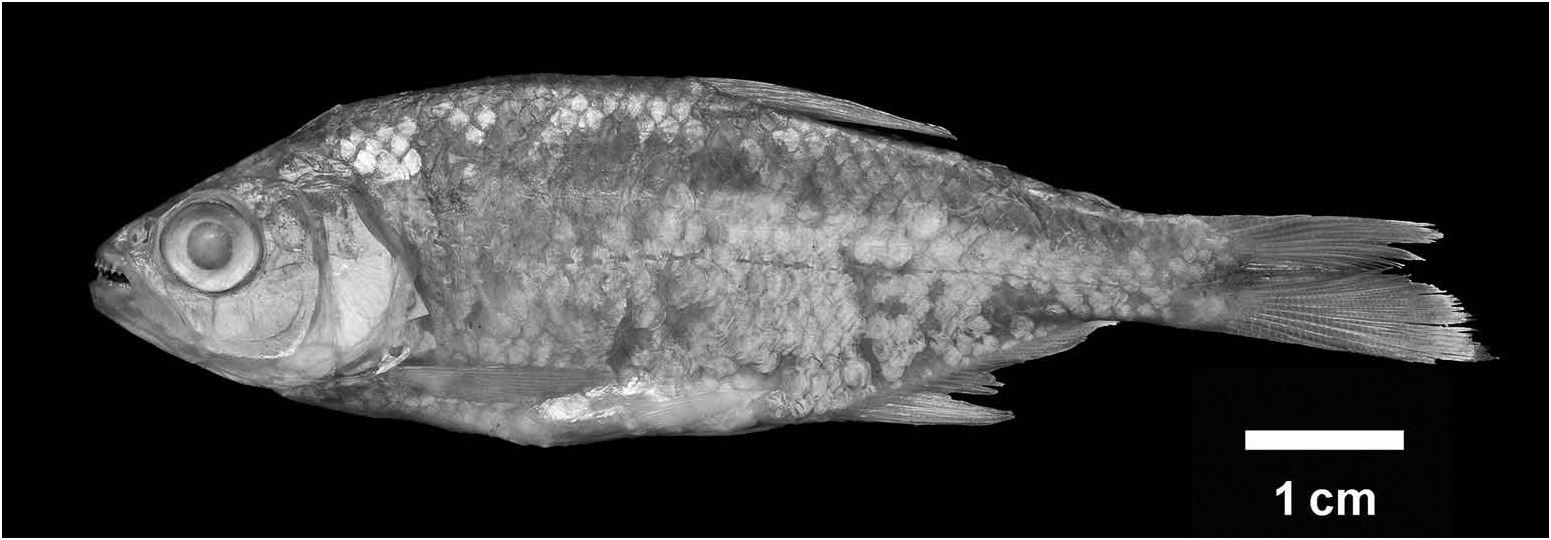
( Figures 8 View Figure 8 (a), 9(a), 15, 18, 19)
Tetragonopterus angustifrons Regan 1908: 172 View in CoL .
Astyanax aeneus View in CoL non Günther, partim. Hubbs 1935 and 1936, Schmitter-Soto et al. 2008, Miller et al. 2009, and others.
Astyanax View in CoL ‘Campeche’, Schmitter-Soto 2016.
Diagnosis
Diagnosed from other Astyanax species in the region as follows: lower-limb gill rakers in first arch, 11–15, mode 13 (9–14, mode 11, in A. fi nitimus; 15–16, in A. ocotal sp. nov.); urohyal, ventrorostral edge convex (angled in A. brevimanus ), its ventral apex closer to the caudal end (about equidistant between rostral and caudal ends in A. fi nitimus); epibranchial III, distal segment straight (curved in A. fi nitimus); preopercle with parallel canals at angle (canals divergent in A. ocotal sp. nov.); coracoid with 2–3 interdigitations in suture to cleithrum (4–5 in A. ocotal sp. nov.); caudal vertebrae, 31 (32–33 in A. brevimanus ); humeral spot, rectangular-triangular (P-shaped in A. brevimanus ).
Redescription
A species of Astyanax , subgenus Astyanax (i.e. with a complete predorsal series of scales).
Head profile, concave to straight-concave; snout round. Lips even; mouth terminal. Pectoral fins usually reach pelvic fin origin; anal and dorsal fins usually do not overlap. Lobes of caudal fin, subequal.
D. 9–11; A. 21–27, modally 24; pect. 9–11. Procurrent unsegmented dorsal rays on caudal fin, variable. Gill rakers on first arch, 20–27, modally 22; on lower arm, 11–15, modally 13. Scales on lateral line, 32–37, modally 33; predorsal scales, 9–13, modally 11; scale rows from lateral line to base of first dorsal-fin ray, 7–8.5, modally 8; to base of pelvic fin, modally 6, up to 7; to base of pectoral fin, modally 4, up to 5; circumpeduncular scales, 14–19, modally 16. A single, short scale row on anal fin base. Nuptial tubercles not observed. Total vertebrae 31–33, 18–19 caudal. Detailed frequencies are given in Table 3.
Largest examined specimen, 98.4 mm SL. Body depth, 27–40% SL. Head length, 22–37% SL; orbital diameter, 27–40% HL; interorbital distance, 7.7–10.4% SL, mean 9.1% SL (further morphometric data appear in Table 4).
Anterior fontanel long, straight-sided, blunt-tipped. Supraoccipital process in dorsal view, variable in length; angled to concave in lateral view. Vomer rostrally slightly concave. Arms of premaxilla, subequal (dentigerous longer in Guatemalan populations); 4 premaxillary teeth. Highest tooth on dentary, first, third or fourth; the posterior teeth, variable in size. Maxillary, with a convex anteroinferior edge, 1–3 teeth. Quadrate, dorsal process not expanded (expanded in Guatemalan populations). Metapterygoid, rostral arm longer than ventral; 2 dorsocaudal projections. Infraorbital II, triangular with an angled base; infraorbital III, inferoposteriorly semicircular; infraorbital IV, squarish to rectangular, with a projection; contact between infraorbitals II and III, wide. Urohyal rostral end turned up, its ventrorostral edge convex, its ventral apex closer to caudal end; ceratohyal foramen drop-shaped; rostral vertices of ceratohyal angled, the ventral side undulate to straight. Epibranchial III, insertion of uncinate process round, the distal segment of the main body straight. Upper pharyngeal bones, oval; lower pharyngeal plate single, its caudal side concave. Dorsal side of hyomandibular, convex. Opercular dorsal edge, usually convex; sides of dorsal half, parallel; caudoventrally straight-convex, caudodorsally concave; ventral tip, sharp. Interopercle, posterior edge, straight-convex, with a spine. Preopercular ventral rim, straight; posteroventral edge, curved; 2 canals at angle, parallel. Four or 5 predorsal bony elements, dorsally expanded. Coracoid with 2–3 interdigitations in suture to cleithrum, a concave caudal edge, a single posteroinferior spine. Caudad process of postcleithrum, variable. Proximal edge of pelvic bone, convex. Postanal element, short. Dorsal tip of scaphium, truncate; caudal tip, spiny; dorsoposterior edge, usually slightly concave. Neural spines under dorsal fin, straight. Fifth to seventh caudal vertebra from tail, with a haemal spine displaced caudad. Epuric plate on last neural spine, edge variable.
Humeral spot, rectangular-triangular or indistinct. Pigment on anal fin sparse, uniform or rather concentrated on first rays. Caudal spot both on peduncle and on fin rays. Live colours were reported for the population at Tuxpeña, inland Campeche ( Hubbs 1936, p. 176, as ‘ A. f. aeneus ’): ‘caudal lemon yellow...anal and pelvic fins bright red towards front...a suffusion of red above anal’. Figure 18 View Figure 18 shows the species live in the upper Río Candelaria, Campeche.
Type material and depositor
Lectotype BMNH 1857.7 .31.9, 78.7 mm SL, ‘Mexico’ (most likely the southern Gulf of Mexico versant, see below), coll. A. Sallé, ca. 1850 ( Figure 19 View Figure 19 ; see Remarks ) . Paralectotypes: BMNH 2016.9 . 13.1–2 (2 specimens), same collection data .
Distribution
Mexico and Guatemala, Campeche and Tabasco to northern Petén ( Figure 15 View Figure 15 ) . The collector, Auguste Sallé, worked in Mexico and Central America, including the southern Gulf region ( Günther 1864), before moving to Pacific Oaxaca (see account for A. aeneus ) .
Proposed common names
Campeche tetra, sardinita de Campeche.
Remarks
In spite of its vague type locality, the species has been recognised as distinct by some authors (e.g. Lima et al. 2003, p. 107), mostly because of the high number of gill rakers, surpassed in Mexico only by A . ocotal sp. nov. (q.v.).
Hubbs (1935, p. 6) identified his material from Petén as A. aeneus based solely on the number of anal-fin rays, and dismissed the possibility of their being A. angustifrons because of the gill rakers being ‘fewer than as described’. However, I counted not 17 but at most 15 gill rakers in the types; perhaps Regan (1908) included rudiments. I have found other discrepancies with Regan’ s counts (see account for A. fi nitimus).
Astyanax angustifrons is a part of clades Ic of Ornelas-García et al. (2008) and D of Strecker et al. (2004). The complexity of Petén, with several Astyanax species present, including two formerly in Bramocharax , was genetically verified by Ornelas-García et al. (2008). See Discussion.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Astyanax angustifrons ( Regan 1908 )
| Schmitter-Soto, Juan J. 2017 |
Tetragonopterus angustifrons
| Regan CT 1908: 172 |