
TYPHLONECTIDAE, Taylor, 1968
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00838.x |
|
persistent identifier |
https://treatment.plazi.org/id/03DB87B7-FFE4-FFA4-FEFA-90F9FDAC677B |
|
treatment provided by |
Marcus |
|
scientific name |
TYPHLONECTIDAE |
| status |
|
TYPHLONECTIDAE View in CoL ( FIGS 10 View Figure 10 , S19)
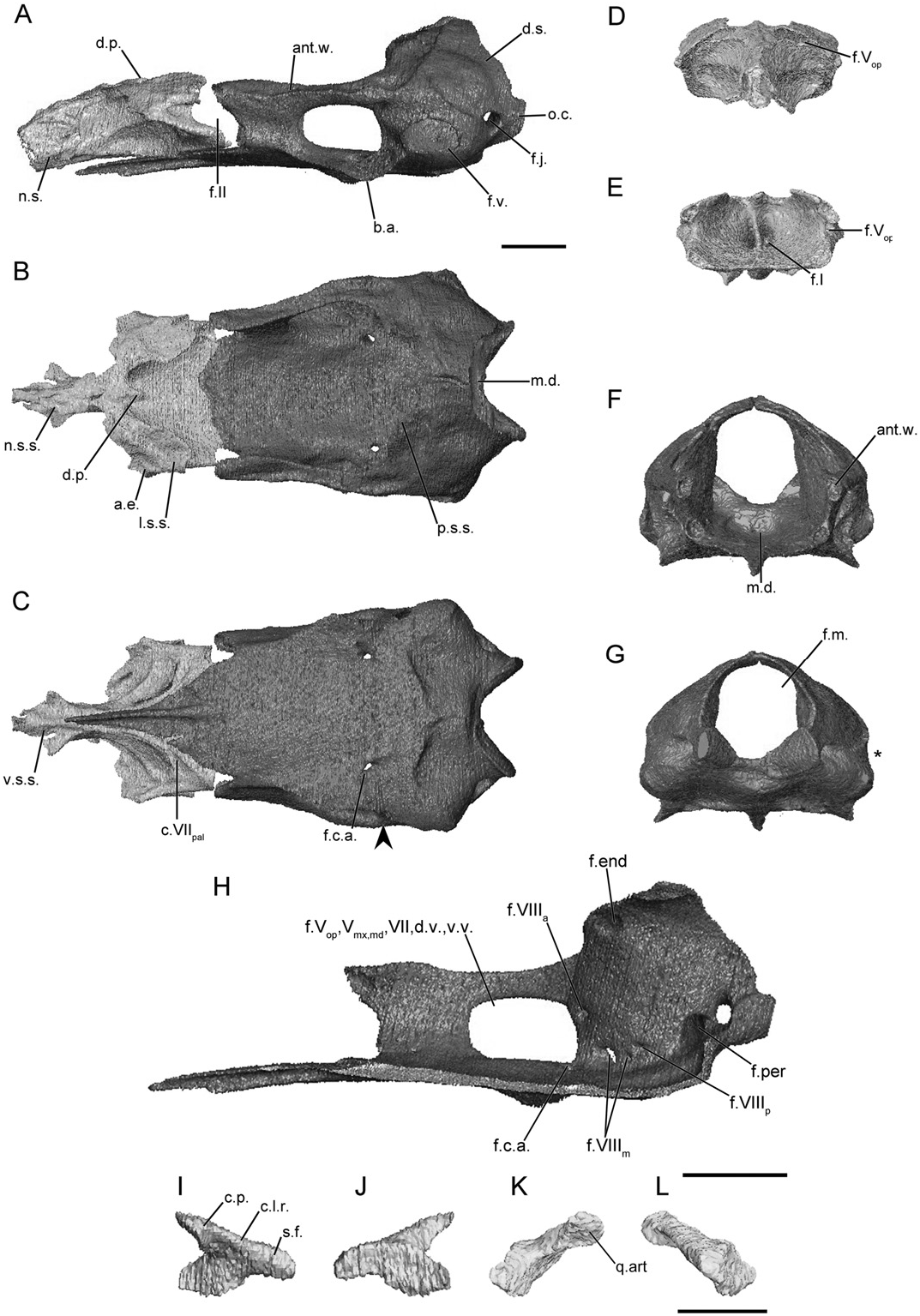
The main body of the sphenethmoid is short and broad in typhlonectids ( Fig. 10A View Figure 10 ). It accounts for roughly half the length of the entire sphenethmoid, which may be more the result of a very short nasal septum region. The lateral wall of the main body is short and bears a very large, broad sutural surface ( Fig. 10B View Figure 10 ), less so in Typhlonectes natans (Fig. S19B). There is an expanded anterolateral corner in Chthonerpeton indistinctum ( Fig. 10B View Figure 10 ) and a thin process in T. natans (Fig. S19B). The dorsomedial process is extremely truncated in Ch. indistinctum , and otherwise short and rod-like in T. natans . In both species it is covered dorsally. The posterior margin of the lateral wall is deeply incised by the anterior margin of the optic foramen ( Fig. 10A View Figure 10 ). The floor of the sphenethmoid is broad and only gently concave in Ch. indistinctum but deeply so in T. natans . No medial incision is present in either species. The ventral surface of the sphenethmoid bears a groove for the palatal ramus of the facial nerve ( Fig. 10C View Figure 10 ).
The nasal septum is very short in typhlonectids, terminating close to the level of the tentacular aperture, rather than at the external naris as it does in most species examined here. It is covered dorsally by a broad sutural surface in Ch. indistinctum ( Fig. 10B View Figure 10 ), and by a narrower surface in T. natans (Fig. S19B). The ventral margin bears a broader sutural surface, and a pair of thin sheet-like processes extends anterolaterally from the surface, in Ch. indistinctum ( Fig. 10C View Figure 10 ). No sola nasi are present in typhlonectids.
The anterior foramina of Ch. indistinctum exhibit a unique configuration amongst the species examined here. The dorsal and ventral foramina on each side have converged to form a single left and single right foramen for the branches of the olfactory nerve ( Fig. 10D, E View Figure 10 ). Paired dorsal and ventral foramina are present in T. natans . There the ventral pair is large and close to the midline (Fig. S19D, E). The anterolateral foramina are present in the bases of the anterolateral corners of the main body ( Fig. 10E View Figure 10 ).
The antotic wall of the os basale is angled anteromedially when viewed dorsally ( Fig. 10B View Figure 10 ). In anterior view it is slightly dorsolaterally orientated in Ch. indistinctum ( Fig. 10F View Figure 10 ) and more vertical in T. natans (Fig. S19F). The dorsal sutural surface is moderately broad, and is continuous with the sutural surface on the anterior margin of the otic capsule ( Fig. 10B View Figure 10 ). The anterior margin of the antotic wall is slightly incised by the posterior margin of the somewhat oblique optic foramen ( Fig. 10A View Figure 10 ). A single, very large foramen is present in the antotic wall of Ch. indistinctum ( Fig. 10H View Figure 10 ), similar to the condition seen in some indotyphlid species (referred to here as Pattern 8; Maddin, 2011). In T. natans the pattern of foramina (Fig. S19H) is like that of caeciliids (referred to here as Pattern 6).
The dorsal surface of the otic-occipital complex of the os basale is not as steeply inclined posteroventrally as it is in caeciliids. It tapers in width towards the midline ( Fig. 10B View Figure 10 ), and a broad portion of the dorsal surface is exposed dorsally at the midline. The sutural surface that receives the parietal tapers in width towards the midline. The fenestra vestibuli is small, and in one specimen ( MW 23 ) has a unique bilobed outline. The fenestra vestibuli does not incise the lateral margin of the otic capsule when viewed posteriorly ( Fig. 10G View Figure 10 ). The occipital condyle does not project far posteriorly ( Fig. 10A View Figure 10 ) and the jugular foramen is visible in lateral view .
The medial wall of the otic capsule is perforated by four foramina in the common locations (endo- and perilymphatic foramina, anterior and posterior vestibulocochlear nerve foramina) found in all species examined here ( Fig. 10H View Figure 10 ). Variably within the family two or three foramina transmitting components of the medial branch of the vestibulocochlear nerve are present ( Fig. 10H View Figure 10 ).
The anterior portion of the floor of the os basale tapers dramatically to a very narrow point ( Fig. 10C View Figure 10 ). The floor extends to reach the ventral surface of the nasal septum. The posterior depression is also weakly defined ( Fig. 10B, F View Figure 10 ). The basicranial articulation is weakly developed in Ch. indistinctum ( Fig. 10A View Figure 10 ), slightly more so in T. natans (Fig. S19A). In ventral view there is only a slight constriction of the lateral margin of the floor posterior to the basicranial articulation. A ventral wing-like projection is absent from the ventral surface of the otic capsule ( Fig. 10A View Figure 10 ). The posterior margin of the floor terminates at a pointed transverse boundary that does not closely approach the ventral margin of the foramen magnum ( Fig. 10C View Figure 10 ).
The foramen in the floor of the os basale that leads to a canal for the carotid artery enters the floor just anterior to the otic capsule and passes out almost directly into the lateral and medial foramina, both of which are located posterior to the large antotic foramen, on either side of the antotic wall ( Fig. 10H View Figure 10 ).
The stapes of typhlonectids is small and gracile in comparison to that of most species. The long axis of the footplate is orientated slightly anteroventrally. The anterior margin of the footplate does not intimately articulate with the margin of the fenestra vestibuli, but it is embedded in dense connective tissue. The columellar process is depressed and anterodorsally orientated ( Fig. 10K View Figure 10 ). There is no foramen in the base of the columellar process ( Fig. 10L View Figure 10 ).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |