
Cnemaspis sundainsula, Grismer & Wood & Anuar & Riyanto & Ahmad & Muin & Sumontha & Grismer & Onn & Quah & Pauwels, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3880.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:03A6448A-25D7-46AF-B8C6-CB150265D73D |
|
DOI |
https://doi.org/10.5281/zenodo.5708554 |
|
persistent identifier |
https://treatment.plazi.org/id/03FA0350-FFC9-2564-FF51-CF3AFD0B2953 |
|
treatment provided by |
Felipe |
|
scientific name |
Cnemaspis sundainsula |
| status |
sp. nov. |
Cnemaspis sundainsula sp. nov.
Sunda Island Rock Gecko
Fig. 54 View FIGURE 54
Gonatodes kendallii Günther 1895:500 ; De Rooij 1915:26 (in part)
Cnemaspis cf. nigridia Leong, Grismer & Mumpuni 2003:170
Cnemaspis kendallii Das & Bauer 1998:13
Holotype. Adult male MZB. Lace. 9438 collected by Awal Riyanto on 24 October 2011 from Mount Ranai , Bunguran Timur district , Natuna Regency , Kepulauan Riau Province, Bunguran (Great Natuna) Island, Indonesia (03 o 57’24.5”N, 108 o 21’08.3”E) at 345 m above sea level GoogleMaps .
Paratypes. Adult male paratypes MZB. Lace 9436–37 and adult female paratypes MZB. Lace 9439–40 have the same data as the holotype except that MZB. Lace 9440 was collected on 25 October 2011; adult maleTNHC 64276, adult female TNHC 62277 View Materials and adult male MZB. Lace. 4621 were collected on 3 April 2003 by B. J. Evans, Mohd. Iqbal Setiadi and Gandhi Probowo from Mount Ranai , Bunguran Timur District, Natuna Regency, Kepulauan Riau Province, Bunguran Island, Indonesia (03°57.381’ N, 108°21.319’ E); GoogleMaps adult female USNM28139 View Materials was collected on 2 July1900 by W. L. Abbott from Bunguran (=Pulau Natuna Besar), Kepulauan Riau Province, Indonesia (03°57.381’ N, 108°21.319’ E); adult males GoogleMaps MZB. Lace 10156 and 10159 were collected on 26 August 2013 and 27 August 2013, respectively by Awal Riyanto and Zamri at Ceruk Forest Conserve , Selemam Village , Bungaran Timur Laut District, Natuna Regency, Kepulauan Riau Province, Bunguran Island, Indonesia (03 o 58’31.6” N, 108 o 17’52.2” E) at 51 m above sea level GoogleMaps ; and adult males MZB. Lace 10160–61 were collected on 28 August 2013 by Awal Riyanto and Zamri at Gunung Air Hiu Recreation Area , Ceruk Village , Bungaran Timur Laut District, Natuna Regency, Kepulauan Riau Province, Bunguran Island, Indonesia (03 o 59’03.7” N, 108 o 19’06.0”E) at 117 m above sea level GoogleMaps .
Additional specimens examined. Adult males MZB.Lace 10157–58 collected on 26 August 2013 and 27 August 2013, respectively by Awal Riyanto and Zamri at Ceruk Forest Conserve , Selemam Village , Bungaran Timur Laut District , Natuna Regency, Kepulauan Riau Province, Bunguran Island, Indonesia ; subadult males MZB.Lace 9439–40 and MZB.Lace 10162 were collected on 28 August 2013 by Awal Riyanto and Zamri at Gunung Air Hiu Recreation Area , Ceruk Village , Bungaran Timur Laut District , Natuna Regency, Kepulauan Riau Province, Bunguran Island, Indonesia .
Diagnosis. Maximum SVL 84.5 mm; 8–11 supralabials; 7–10 infralabials; ventral scales keeled; no precloacal pores; 26–37 paravertebral tubercles; tubercles linearly arranged, preent on flanks but absent in lateral caudal furrows; ventrolateral caudal tubercles present; lateral row of caudal tubercles present; caudal tubercles not encircling tail; subcaudals smooth but bearing an enlarged median row of smooth scales occasionally posteriorly; 2–4 postcloacal tubercles on each side of tail base; no enlarged femoral or subtibial scales; submetatarsal scales of first toe weakly enlarged to enlarged; subtibials keeled; 25–29 subdigital fourth toe lamellae; small, light-colored round sponts on flanks; gukar region, throat, and lateral sections of abdomen orange; anterior subcaudal region yellow, posterior region white (Tables 6,7).
Description of holotype. Adult male, SVL 84.1 mm; head robust, oblong in dorsal profile, moderate in size (HL/SVL 0.23), not narrow (HW/SVL 0.15), flattened (HD/HL 0.41), distinct from neck; snout moderate (ES/HL 0.42), slightly concave in lateral profile; postnasal region constricted medially; scales of rostrum keeled, raised, larger than scales on occiput; distinct, supraorbital ridges; no frontorostral sulcus; canthus rostralis rounded; eye large (ED/HL 0.22); extra-brillar fringe scales largest anteriorly; pupil round; ear opening oval, taller than wide; rostral concave dorsally, posterior 90% divided by longitudinal groove; rostral bordered posteriorly by two large supranasals and external nares, laterally by first supralabials; 9R,L raised supralabials decreasing in size posteriorly; 8R,L infralabials, decreasing in size posteriorly; nostrils round, oriented dorsoposteriorly; mental large, triangular, flat, bordered posteriorly by six postmentals, first two on either side largest; gular and throat scales granular, keeled, raised; pectoral scales slightly larger.
Body robust (AG/SVL 0.43); small, granular, rugose, dorsal scales generally equal in size throughout body, intermixed with larger, multicarinate tubercles more or less linearly arranged; tubercles extend from occiput to base of tail; tubercles on flanks; 32 paravertebral tubercles; pectoral and abdominal scales small, granular, keeled, same size throughout; abdominal scales slightly larger than dorsals; no precloacal pores; forelimbs moderately long, robust (FL/SVL 0.19); dorsal scales of brachium raised, keeled; dorsal scales of forearm raised, keeled; ventral scales of brachium keeled, raised, juxtaposed; ventral scales of forearm keeled, raised, juxtaposed; palmar scales, juxtaposed, raised, broadly keeled; digits long with an inflected joint; claws recurved; subdigital lamellae unnotched; lamellae wide throughout digit; interdigital webbing absent; fingers increase in length from first to fourth with fourth longer than fifth; hind limbs robust, slightly longer and thicker than forelimbs (TBL/SVL 0.24); dorsal scales of thigh keeled, raised, juxtaposed; scales of anterior margin of thigh keeled; ventral scales of thigh keeled; subtibial scales raised, keeled, juxtaposed, with no enlarged anterior row; plantar scales smooth, juxtaposed, raised; weakly enlarged to enlarged submetatarsal scales beneath first metatarsal; digits elongate with an inflected joint; claws recurved; subdigital lamellae unnotched, wide throughout digit; interdigital webbing weak to absent; toes increase in length from first to fourth with fourth being slightly longer than fifth; 28 subdigital lamellae on fourth toe; caudal scales arranged in segmented whorls; dorsal caudal scales raised, keeled, juxtaposed; deep middorsal and lateral caudal furrows; subcaudal scales smooth; median row of enlarged subcaudal scales posteriorly; caudal tubercles do not encircle tail; tubercles absent from lateral furrows; three enlarged postcloacal tubercles on lateral surface of hemipenal swellings at base of tail.
Coloration in life ( Fig. 54 View FIGURE 54 ). Dorsal ground color of head, body and limbs yellowish brown; paired, yellowish, lineate markings on rostrum; irregularly shaped yellowish markings on top of head; thin, black, upper postorbital stripe extending to occiput; thin, black, lower postorbital stripe extending onto flank; thin, yellowish, postorbital stripe highlighting row of tubercles on latter surface of occiput; transverse, beige marking on nape followed by one square and two rectangular medial beige markings on body alternating with small, elongate, thin, black vertebral markings; light rectangular markings on body grade into diffuse, light caudal bands alternating with darker brown bands; caudal bands do not encircle tail; diffuse light blotches on flanks; tubercles on flanks white; dark, diffuse, rectangular markings on back and flanks alternate with light markings; gular region and throat yellow-orange; lateral margins of abbomen and lower flanks yellow-orange; anterior one-half of subcaudal region yellow, posterior one-half white; ventral surfaces of pectoral region and limbs beige.
Variation ( Fig. 54 View FIGURE 54 ). All paratypes closely resemble the holotype in coloration and pattern although TNHC 64276–77 and MZB.Lace 4621 are not nearly as boldly marked. The color pattern of MZB.Lace 10156 is much bolder than that of the holotype in that the dark and light dorsal markings stand in distinct contrast to one another. The dark dorsal markings of MZB.Lace 10161 are elongate as opposed to being more square to roundish as in the other specimens. TNHC 64277 lacks a tail and the tail of TNHC 64276 is regenerated and composed of small, dark, roundish, juxtaposed scales that are weakly keeled on the dorsal surface whereas the subcaudals are smooth beige, and slightly larger. USNM 28139 is badly faded and only a general color pattern that matches that of the holotype is visible. Hatchlings tend to have yellow undersides. Meristic differences are listed in Table 12 View TABLE 12 .
Comparisons. Within the Southern Sunda clade, Cnemaspis sundainsula sp. nov. is differentiated from the species of the nigridia group ( C. nigridia and C. paripari ) by having a greater maximum SVL (84.5 mm versus 50.7–75.5 mm); lacking as opposed to having precloacal pores; having tubercles on the flanks as opposed to lacking them; having smooth as opposed to keeled subcaudals; generally lacking as opposed to having a median row of enlarged, subcaudal scales; and lacking as opposed to having weakly enlarged, metatarsal scales beneath the first toe. From the species of the kendallii group ( C. baueri , C. pemanggilensis , C. mumpuniae sp. nov., C. bidongensis , and C. peninsularis sp. nov.) of which it is a member, it can be differentiated by having a much greater maximum SVL (84.5 mm versus 58.1–76.0 mm), having smooth as opposed to keeled subcaudal scales, and not having caudal tubercles that encircle the tail. From C. limi , C. sundainsula sp. nov. is separated by having linearly as opposed to randomly arranged dorsal tubercles, and having a ventrolateral row of caudal tubercles. Cnemaspis sundainsula sp. nov. can be differentiated from C. sundagekko sp. nov, by having a much larger maximum SVL (84.5 mm versus 68.0 mm), a greater number of paravertebral tubercles (26–37 versus 20–25), smooth as opposed to keeled subcaudals, caudal tubercles that do not encircle the tail, and fewer subdigital lamellae on the fourth toe (26–31 versus 33–38).
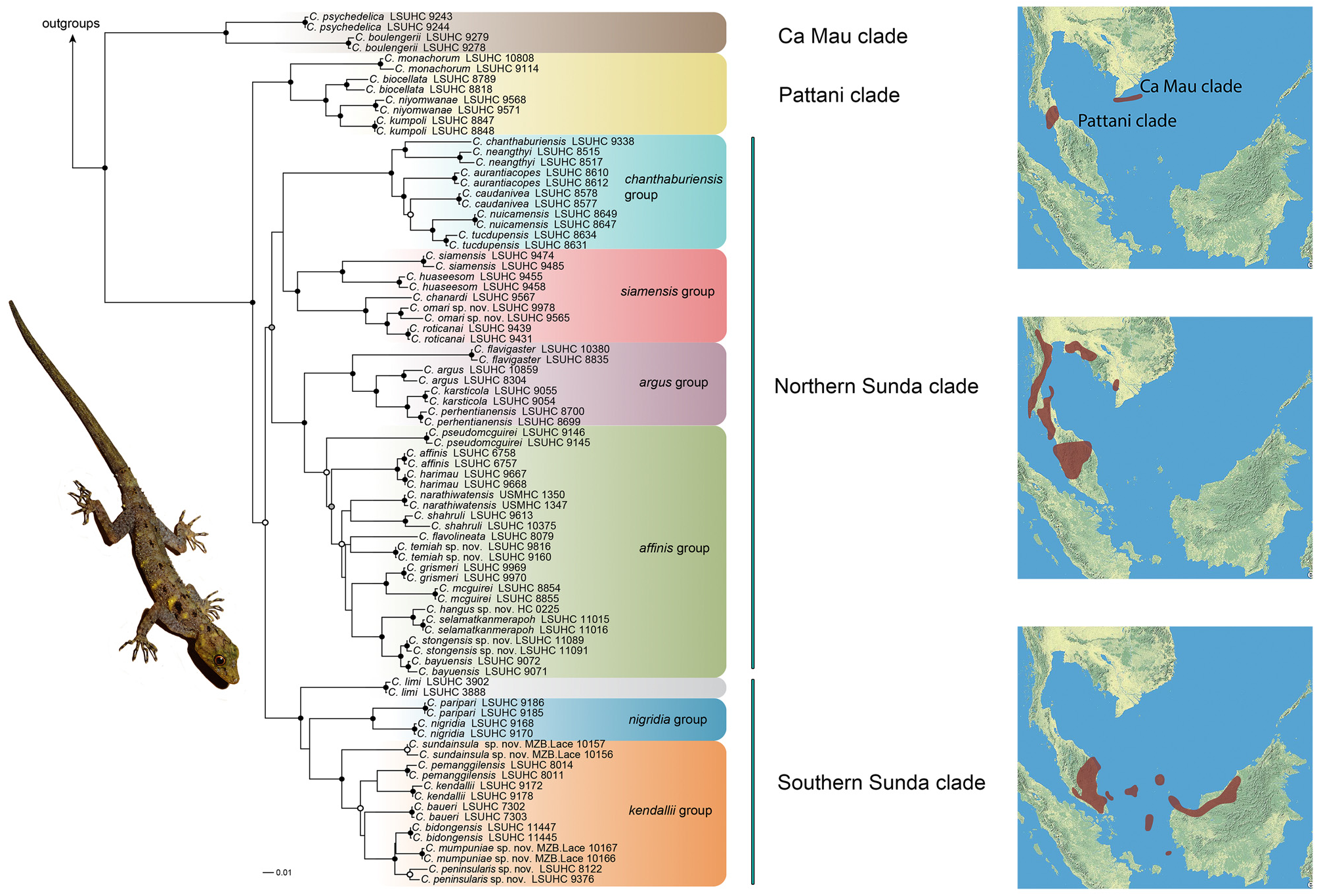
Distribution. Cnemaspis sundainsula sp. nov. is known only from Bunguran Island (= Pulau Natuna Besar/ Great Natuna) of the Natuna Archipelago, Kepulauan Riau Province, Indonesia ( Fig. 4 View FIGURE 4 ) but is likely to occur on nearby islands in the archipelago.
Natural History. Cnemaspis sundainsula sp. nov. inhabits primary and secondary forests along the base of Mount Ranai from at least 51–345 m above sea level ( De Rooij 1915; Günther 1895). It is not certain if this species extends up to the summit but if granite boulders are present we suspect it does. Lizards occur almost exclusively on large granite boulders ( Fig. 54 View FIGURE 54 ) and are only rarely found on tree trunks. This species is abundant and active during the day while in its dark color phase ( Fig. 54 View FIGURE 54 ) and often in male-female pairs. While active during the day, lizards remain wary and will not venture too far out onto the exposed boulder surfaces but rather remain near cracks or narrow spaces between adjacent boulders were escape is possible. Lizards are commonly seen upside down suspended from the undersides of boulders with their limbs outstretched displaying their birght-orange lower flanks, throat, and abdomens ( Fig. 54 View FIGURE 54 ). Before escaping into cover lizards, roll their tails up over their backs and display the immaculate, white posterior subcaudal region ( Fig. 54 View FIGURE 54 ). In dark crevices and boulder spaces, often the subcaudal region is all that can be seen. At night, lizards venture farther out onto the exposed boulder surface and sequester themselves in a shallow depression or crease or along the edge of a small ridge where they are generally inactive. During this period, lizards are in their light color phase ( Fig. 54 View FIGURE 54 ) and are much more approachable. We have observed gravid females carrying two eggs, hatchings, and juvenlies during April.
Cnemaspis sundainsula sp. nov. occurs syntopically with C. mumpuniae sp. nov. and Cyrtodactylus hikidai Riyanto when the latter two species occur on granite boulders. The ecological pattern of temporal partitioning on islands seen between C. sundainsula sp. nov. and Cyrtodactylus hikidai is similar to that between C. psychedelica on Hon Khoai Island, Vietnam which is diurnal and found syntopically with the nocturnal Cyrtodactylus sp. nov. 1 ( Grismer et al. 2010b); C. boulengerii on Con Son Island in the Con Dao Archipelago, Vietnam which is diurnal and syntopic with the larger Cyrtodactylus condorensis (Smith) ; and the large, diurnal, granite boulder-dwelling C. limi from Pulau Tioman, Peninsular Malaysia which is syntopic with the larger nocturnal Cyrtodactylus tiomanensis Das & Lim. Additionally , C. sundainsula sp. nov. and C. mumpuniae sp. nov. partition their habitat by SVL, elevation, and general microhabitat preference where C. sundainsula sp. nov. is a large, rock-dweller and C. mumpuniae is the smaller, habitat generalist. A parallel system exists on Pulau Tioman, Malaysia with the smaller, habitat generalist C. peninsularis sp. nov. and the larger, rock-dwelling C. limi ( Grismer 2011a) .
Etymology. The specific epithet sundainsula is derived from the word Sunda which originally referred to a Hindu Kingdom in western Java existing from 669–1579. Sunda is now commonly used as an adjective associated with particular geographic features in the western regions of Southeast Asia associated with the South China Sea and its fringing continental areas. The Latin insula (singular) means island and sundainsula is an invariable noun in apposition in reference to this species being endemic to an island on the submerged Sunda Plains.
Relationships. Cnemaspis sundainsula sp. nov. is part of the kendallii group within the Southern Sunda clade and is basal to a group containing C. pemanggilensis , C. kendallii sensu stricto, C. baueri , C. mumpuniae sp. nov., C. bidongensis , and C. peninsularis sp. nov. ( Fig. 2 View FIGURE 2 ).


| MZB |
Museum Zoologicum Bogoriense |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cnemaspis sundainsula
| Grismer, Lee, Wood, Perry L., Anuar, Shahrul, Riyanto, Awal, Ahmad, Norhayati, Muin, Mohd A., Sumontha, Montri, Grismer, Jesse L., Onn, Chan Kin, Quah, Evan S. H. & Pauwels, Olivier S. A. 2014 |
Cnemaspis cf. nigridia
| Leong, T. - M. & Grismer, L. L. & Mumpuni 2003: 170 |
Cnemaspis kendallii
| Bauer, A. M. & Das, I. 1998: 13 |
Gonatodes kendallii Günther 1895:500
| De Rooij, N. 1915: 26 |
| Gunther, A. 1895: 500 |