
Conus, Linnaeus, 1758
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2003.00058.x |
|
persistent identifier |
https://treatment.plazi.org/id/03B6B923-EE17-FFC1-8FF1-609639E86873 |
|
treatment provided by |
Carolina |
|
scientific name |
Conus |
| status |
|
CONUS View in CoL ( LEPTOCONUS ) JASPIDEUS GMELIN 1791
Material Examined
Little Duck Key, Florida ( USNM 890945). Only immature males were available; female anatomy is unknown, and male reproductive anatomy is preliminary.
External anatomy and mantle cavity
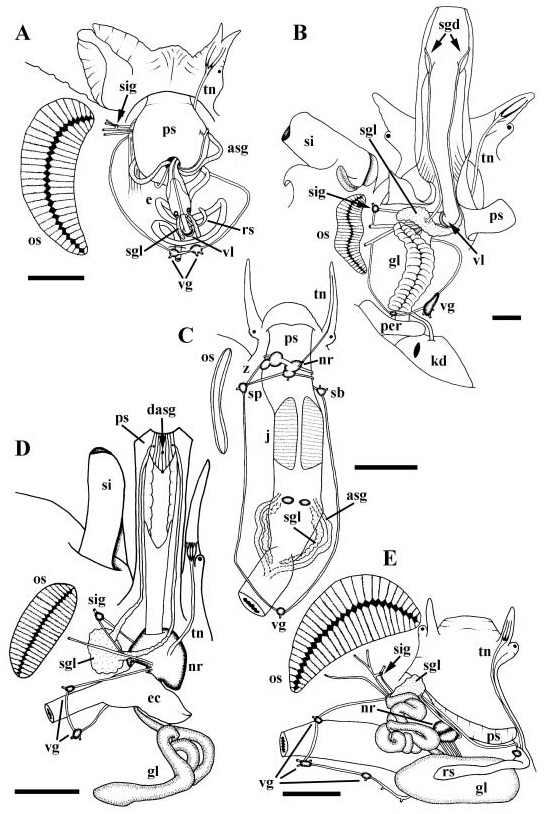
Mantle edge scalloped. Siphon short and broad with asymmetrical basal flaps. Left flap large, lying against side of head. Ctenidium and large bipectinate osphradium ( Fig. 7E View Figure 7 , os) lying short distance behind base of siphon. Large, folded hypobranchial gland present. Anterior pedal gland opening to slit under thick propodial flap. Ventral pedal gland unknown. Operculum present.
Reproductive system
Gonad occupying tip of viscera, posteriorly bordering digestive gland. Vas deferens emerging from testis, rapidly developing into highly convoluted seminal vesicle. Prostate opening to mantle cavity via proximal slit. Closed pallial vas deferens traversing mantle floor and side of neck to penis behind right cephalic tentacle. Penial duct opening at tip of penis.
Alimentary system
Foregut. Proboscis intraembolic ( Fig. 7E View Figure 7 , ps). Rhynchostome with sphincter opening to spacious rhynchocoel. Tentacles surrounding rhynchostome absent. Entire surface of rhynchocoel lined with tall, glandular, basophilic folds; discrete snout gland absent. Proboscis wall folded upon itself where sheath attaches to head wall. Proboscis sheath extending only short distance beyond base of rhynchocoel. Approximately six to eight large retractor muscles extending from base of proboscis, to wall of cephalic haemocoel. Proboscis telescopically folded within rhynchocoel, with true mouth at tip, and intermediate sphincter short distance behind mouth. Single tooth present within proboscis tip. Buccal lips surrounding slit-like opening at tip. Short buccal tube leading to muscular buccal mass lying at base of proboscis; odontophore absent. Radular sac (rs) and poison gland duct (gl) entering buccal mass on right side, latter just posterior to former. Radular sac (rs) subdivided into long and short arms. Posterior, long arm secreting radular teeth, connecting to radular caecum via small constriction. Radular caecum small, lined with cuticle, storing numerous mature teeth within lumen. Paired salivary gland ducts extending from mass comprising paired ascinous salivary gland (sgl), around anterior oesophagus, one on each side, opening to caecum. Long, narrow, glandular poison gland duct, opening to large, muscular bulb (gl). Accessory salivary gland absent. Short, broad anterior oesophagus uniformly folded ( Fig. 10E View Figure 10 ).
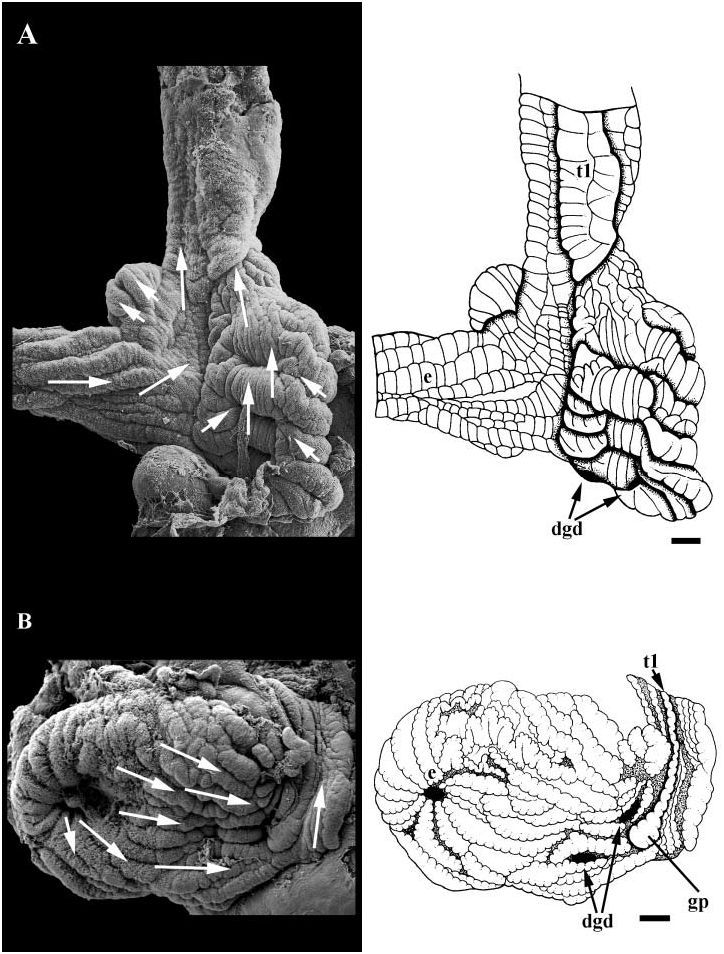
Midgut. Midgut small, U-shaped, embedded in digestive gland just posterior and to left of kidney. Lumen lined with longitudinal, uniformly ciliated folds ( Fig. 18B View Figure 18 ). Paired digestive gland ducts present (dgd). One duct draining posterior lobe, opening to back wall on right. Second duct draining anterior lobe, opening near intestinal aperture. Major typhlosole (t1) extending posteriorly into gastric chamber, proximal tip expanding into large pad near anterior most digestive gland duct. Style sac region bearing paired, glandular typhlosoles and longitudinally folded epithelium bearing undifferentiated cilia. Ciliary currents flowing linearly within gastric chamber from oesophageal aperture to style sac region.
Hindgut. Intestine straight. Anal gland forming narrow, muscular, unbranched tubule, opening to rectum short distance behind anus.
Reno-pericardial system
Kidney long. Afferent renal vessel entering kidney floor anteriorly, adjacent to nephropore and reno-pericardial canal. VARV extending posteriorly within floor, supplying primary tubules along right wall and roof. DARV extending postero-dorsally, running along right margin of roof, supplying secondary tubules within roof. Primary and secondary tubules pycnonephridial, interdigitating within kidney roof along narrow band extending approximately three-quarters length of kidney; primary tubules exclusively lining posterior onequarter of kidney roof. Nephridial gland present.
Nervous system and sensory structures
Nervous system epiathroid, right zygoneurous, and left dialyneurous. Nerve ring lying obliquely within cephalic haemocoel near base of proboscis ( Fig. 7E View Figure 7 , nr). Single nerve branching near cephalic tentacle, giving rise to tentacular and optic nerves. Single tentacular nerve splitting rapidly into three nerves of equal size near eye (tn). Long buccal connectives innervating buccal ganglia lying between anterior oesophagus and radular caecum. Double commissure present between buccal ganglia. Pedal and suboesophageal connectives long, extending around spacious anterior oesophagus. Supra-oesophageal ganglion lying near nerve ring innervated by short connective. Small siphonal ganglion (sig) present at point of dialyneury between branch of anterior osphradial nerve and pallial nerve. Single stout siphonal nerve emerging from ganglion. Pedal ganglia large with propodial ganglia lying at their anterior bases. Visceral loop bearing three visceral ganglia (vg). Large statocysts with single, large statoliths present ventrally and anterior to pedal ganglia.
Discussion
The pallial cavity of conids is dominated by a long and complex bipectinate osphradium ( Taki, 1937; Alexander, 1970, 1973; Alexander & Weldon, 1975; Taylor & Miller, 1989). Size and orientation of the structure and complexity of individual leaflets has been shown to vary. Several variables, including curvature and density of leaflets showed no correlation with ecological factors suggesting a possible utility in phylogenetic studies ( Spengler & Kohn, 1995).
There exists a rather large body of comparative data on conid radular morphology (e.g. Nybakken, 1970a,b, 1990; James, 1980; Shimek & Kohn, 1981; Kohn, Nishi & Pernet, 1999), particularly in the context of feeding ecology. Indeed, shifts in feeding have occurred rarely within Conus , with radular types cutting across patterns of feeding biology. This indicates that, in general, radular tooth morphology is a rather reliable indicator of phylogeny ( Duda, Kohn & Palumbi, 2001).
Prososcis structure is rather uniform in the Conidae compared to other families in the Conoidea (e.g. Miller, 1989), and comprises an intraembolic proboscis with a buccal mass at its base; the valve of Leiblein is lacking. A poison gland consisting of a large, muscular bulb and a long, glandular duct, opens to the buccal mass. Radular teeth are formed in the long arm of the radular sac and passed to the short arm for storage before being transferred to the proboscis tip. Much of the variability of foregut anatomy concerns the accessory glands. A snout gland that opens to the rhynchocoel may be present or absent; accessory salivary glands may be present or absent, single or paired (e.g. Bergh, 1895b; Amaudrut, 1898; Shaw, 1914; Alpers, 1931; Jaeckel, 1952; Abbott, 1967; Marsh, 1971, 1977; Greene & Kohn, 1989; Miller, 1989). Conus jaspideus corresponds to Feeding Mechanism 4 in the classification scheme of Kantor (1990). Several comparative surveys of neogastropod and/or toxoglossan foregut anatomy also are available ( Taylor & Morris, 1988; Miller, 1989; Kantor & Taylor, 1991; Medinskaya, 1992; Kantor, 1996); Taylor, Kantor & Sysoev (1993) provided a review of foregut structures within the Conoidea and a revised classification of the Conidae based largely on this system.
The midgut of conids is small and undifferentiated, lacking caecae and sorting areas ( Shaw, 1914). A discussion of the presence or absence of the anal gland within the Conidae is a notable exception in discussions of neogastropod phylogeny, despite being commonly reduced or lost in many neogastropods (e.g. Ponder, 1973; Taylor & Morris, 1988); Shaw (1914) did not figure or describe an anal gland in Conus tulipa or C. textile . There are no descriptions of renal morphology within the Conidae .
Available descriptions converge on a consistent view of the nervous system as zygoneurous, dialyneurous, with long subesophageal, buccal and pedal connectives. The supra-oesophageal ganglion is incorporated in the nerve ring and there are three visceral ganglia along the visceral loop. The tentacular nerve gives rise to the optic nerve, then branches into three nerves of equal size. The statocysts lie ventral to the circumoesophageal nerve ring ( Bouvier, 1887; Shaw, 1914).

| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |