
Cuthonella concinna, (Alder & Hancock, 1843)
|
publication ID |
https://doi.org/ 10.1163/18759866-BJA10017 |
|
persistent identifier |
https://treatment.plazi.org/id/038C8782-E47D-924C-FF7E-FF3AFB654C3A |
|
treatment provided by |
Felipe |
|
scientific name |
Cuthonella concinna |
| status |
|
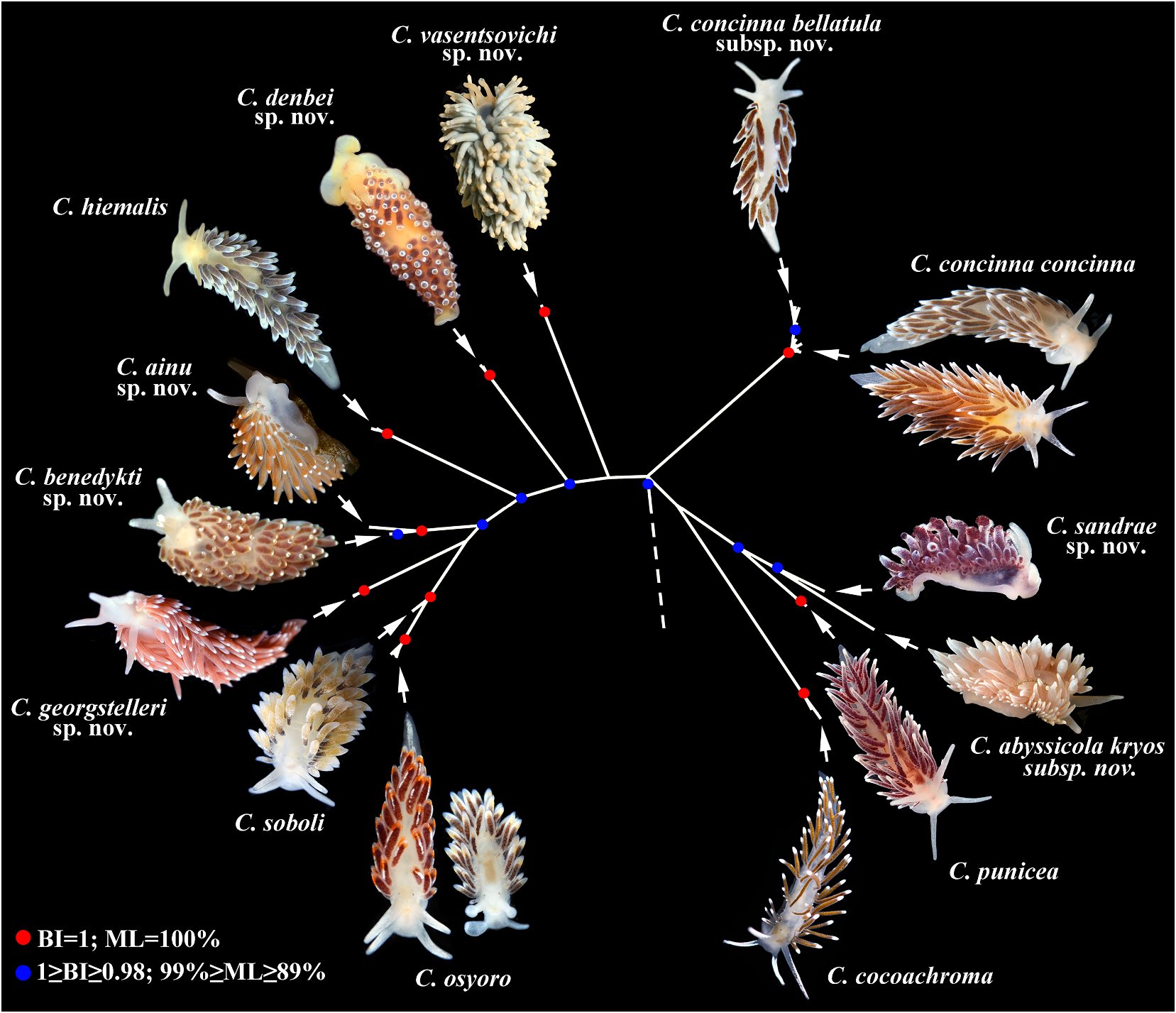
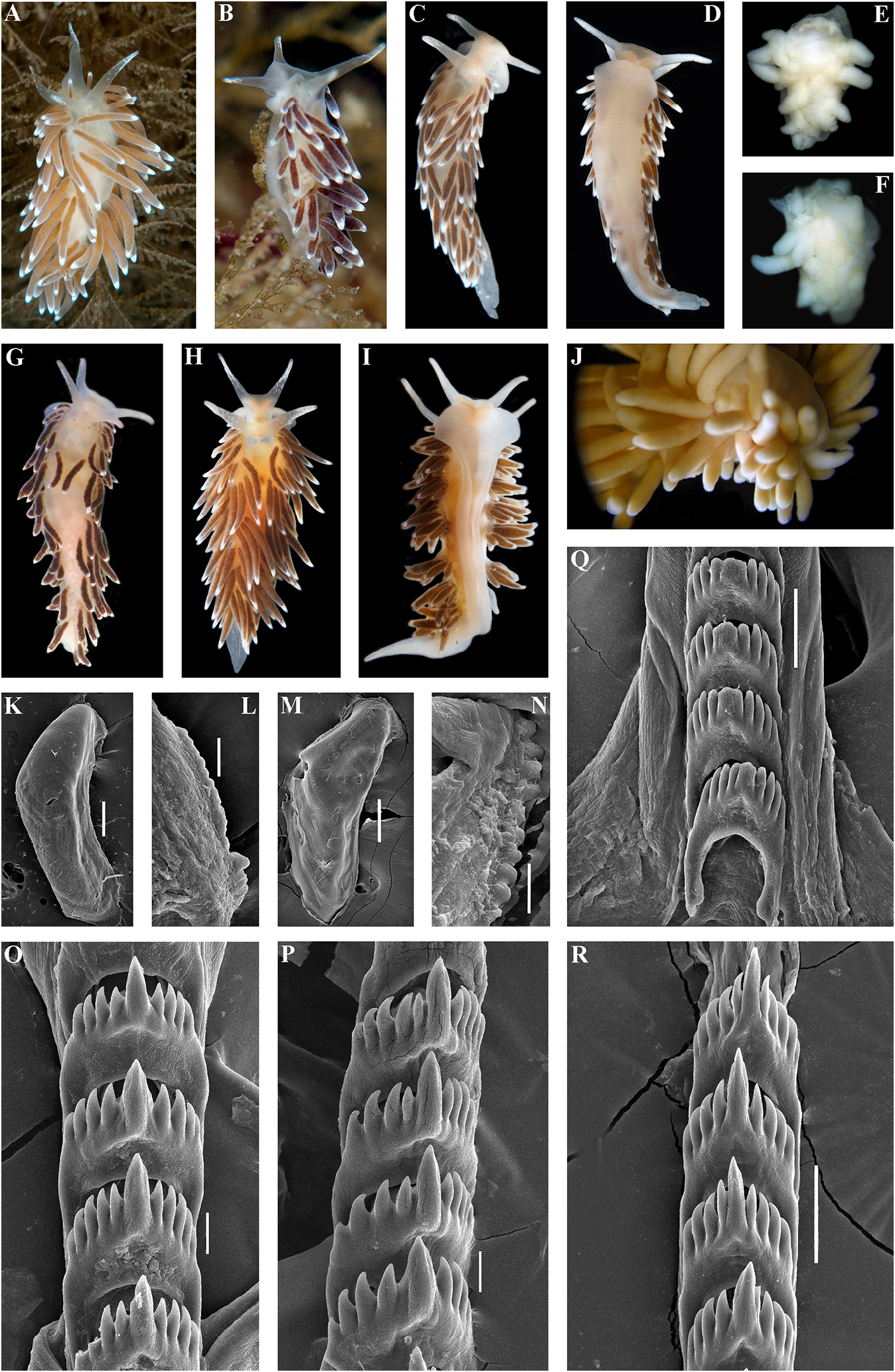
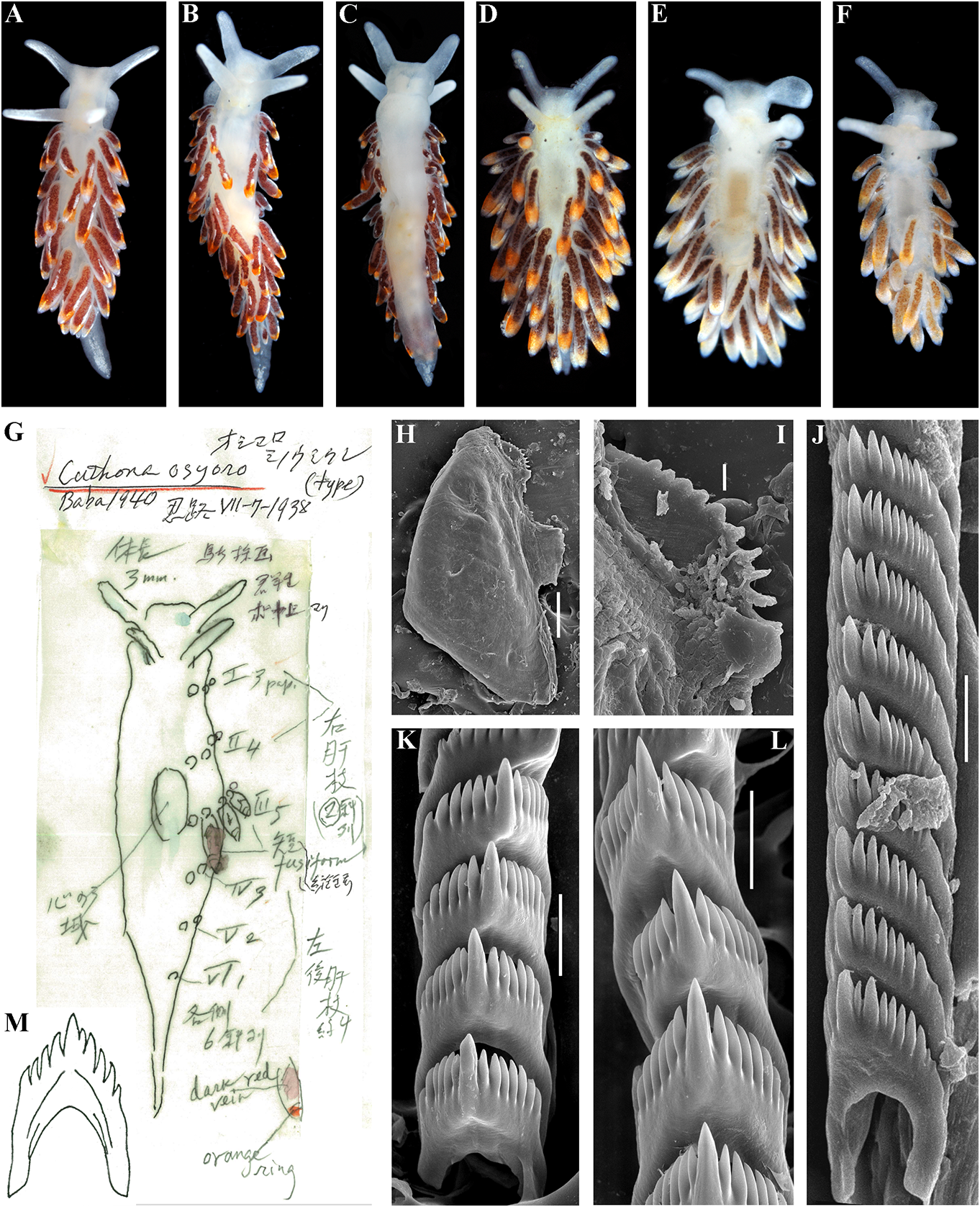
Systematic account concinna (Alder & Hancock, 1843) View in CoL (fig. 8), C.
Family Cuthonellidae Miller, 1977 concinna bellatula subsp. nov. (fig.9), C.denbei Diagnosis. Aeolidaceans with a wide to sp. nov. (fig. 10), C. elenae Martynov, 2000 , moderately broad body. Notal edges fully C. georgstelleri sp. nov. (fig. 11), C. hiemalis reduced. Cerata non elevated, several per row. ( Roginskaya, 1987) (fig. 12), C. orientosiberica Ceratal rows branched or simple.Rhinophores sp. nov. (fig. 13), C. osyoro (Baba, 1940) (fig. 14), smooth to wrinkled, without distinct lamellae. C. punicea (Millen, 1986) (fig. 15), C. sandrae Anus acleioproctic or cleioproctic, in anterior sp. nov. (fig. 16), C. soboli Martynov, 1992 (fig.
to middle part of dorsal side. Radula formula 17), C. vasentsovichi sp. nov. (fig. 18).
0.1.0. Rachidian teeth usually with strong cusp, Remarks. Until1992,the genus Cuthonella was not compressed by adjacent lateral denticles. rarely used to refer just to the deep-sea species, Distal receptaculum seminis present. Vas C. abyssicola , and its two synonymous taxa (e.g., deferens usually moderate, with indistinct Odhner in Franc, 1968; Miller, 1977), or more prostate. Massive external permanent penial often it was considered invalid (Millen, 1986).
collar absent. Penis internal, unarmed. Other species that demonstrate morphological Supplementary gland present, commonly similarities to the type species C. abyssicola inserts to vas deferens. were never considered as belonging to the genus Type genus Cuthonella Bergh, 1884 . Cuthonella infaunallistsandguides(e.g.,Odhner, 1907, 1939; Thompson & Brown, 1984; Behrens, Genus Cuthonella Bergh, 1884 1980; Behrens & Hermosillo, 2005). Martynov ( Figs 1–22 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 View FIGURE 18 View FIGURE 19 View FIGURE 20 View FIGURE 21 View FIGURE 22 ) (1992) initially revised the composition of the Cuthonella Bergh, 1884: 23–24 . genus Cuthonella to include six species – C.
Cuthonella Martynov, 1992 View in CoL , revision: 23. abyssicola (Bergh, 1884) View in CoL , C. concinna View in CoL (Alder Non Cuthona sensu Millen (1986) View in CoL & Hancock, 1843), C. distans ( Odhner, 1922) , Type species: Cuthonella abyssicola Bergh View in CoL , C. hiemalis ( Roginskaya, 1987) View in CoL , C. marisalbi View in CoL 1884. Recent, North Atlantic Ocean. Gender (Roginskaya, 1963), C.punicea (Millen, 1986) View in CoL and feminine. to describe a new one, C.soboli Martynov, 1992 View in CoL .
Diagnosis. Body wide to moderately broad Thus, a future monophyletic molecular (rarely more narrow). Cerata numerous, clade (fig. 1) was predicted using only continuous. Rhinophores smooth to wrinkled, morphological data. This is a clear indication longer or similar in size to oral tentacles. that morphological data (ontogenetic in a Anterior foot corners never tentaculate. broad sense) should have value independent Anus acleioproctic (rarely cleioproctic). of molecular data (Korshunova et al., 2017a, b) Central teeth with strong cusp and distinct instead of being considered merely an inferior denticles. Distal receptaculum seminis. Long auxiliary addition, a view commonly held in vas deferens without separate granulated the current use of molecular techniques to prostate. Supplementary gland (= “penial delineate species (e.g., Carmona et al., 2013).
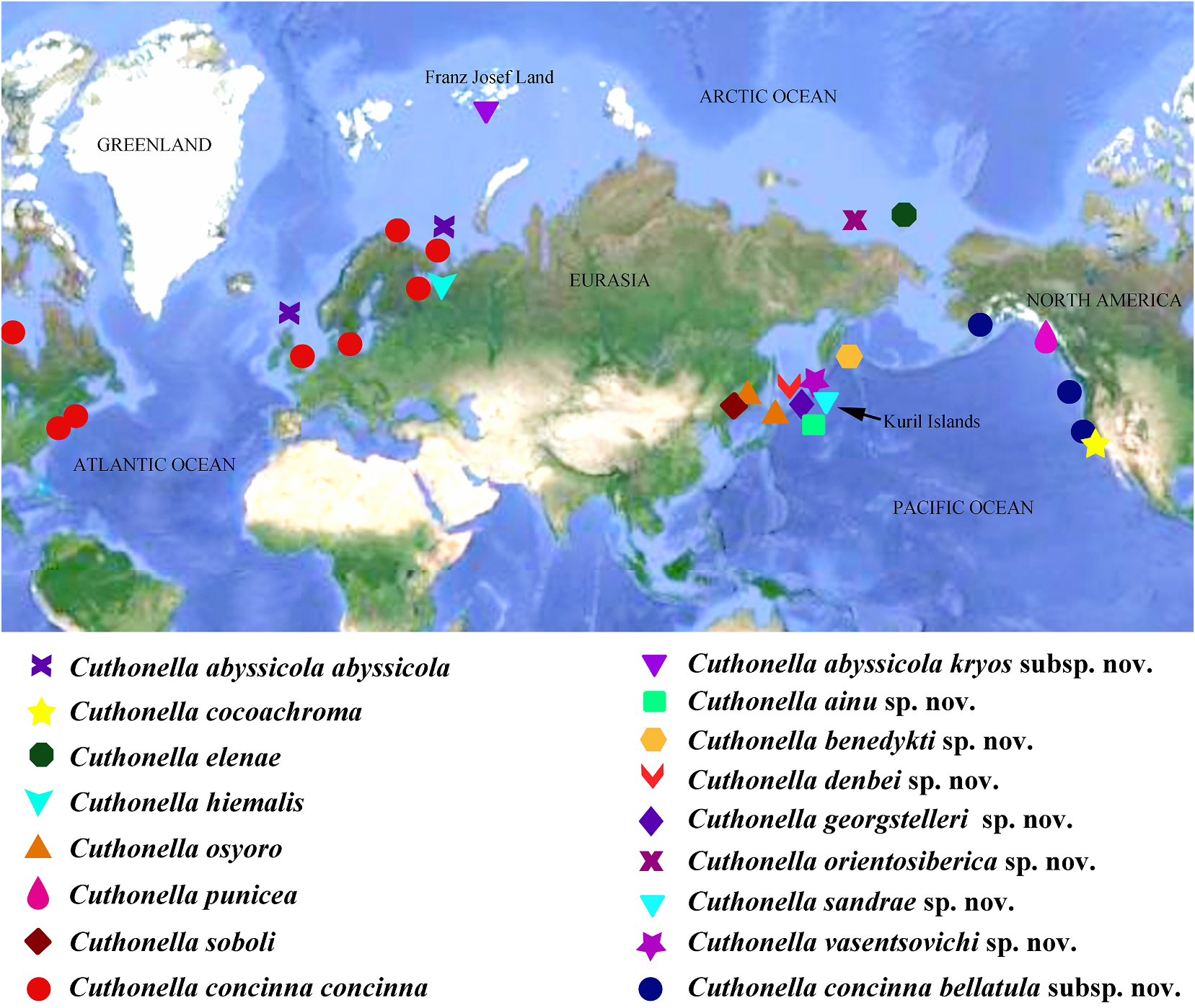
gland”) inserts into vas deferens. Penis In the present study, five of the seven commonly elongated conical, unarmed. species that were included in the genus Contained species: Cuthonella abyssicola Cuthonella by Martynov (1992) are confirmed abyssicola Bergh, 1884 (fig. 4), C. abyssicola to belong to the monophyletic molecular kryos subsp. nov. (fig. 5), C. ainu sp. nov. (fig. clade that includes Cuthonella abyssicola s.l.
6), C. benedykti sp nov. (fig. 7), C. cocoachroma Moreover , Cuthonella concinna s.l., C. hiemalis ( Williams & Gosliner, 1979) , C. concinna (Roginskaya Downloaded, 1987from), C. Brill punicea .com 12(/ Millen 12/2023, 04 1986:16:22) PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/
and C. soboli Martynov, 1992 View in CoL are confirmed denticles is well attested for the type species to be valid species (figs 1–3, 4, 8, 12, 15, 17), of Guyvalvoria View in CoL , G. francaisi Vayssière, 1906 View in CoL whereas C. distans ( Odhner, 1922) and C. with similar body size (up to 14.5 mm marisalbi (Roginskaya, 1963) View in CoL – while still preserved length), whereas another Antarctic Cuthonella View in CoL as predicted by Martynov (1992) – species Guyvalvoria gruzovi Martynov, 2006 View in CoL
are now both considered to be junior has a clearly larger number (11–15) of less synonyms of Cuthonella concinna concinna View in CoL distinct lateral denticles in specimens with
(see below and fig. 8). smaller (8–9 mm) preserved body length Previously, a few Antarctic species were ( Martynov , 2006a: 78–84). In addition, described under the genus Cuthonella , i.e. Cuthona schraderi (Pfeffer in Martens & Cuthonella antarctica Eliot, 1907 (currently is Pfeffer, 1886) was considered as a potential known under a replacement name Cuthonella synonym of G. “paradoxa ” sensu Valdés et al.
elioti (Odhner, 1944)) View in CoL , Cuthonella modesta View in CoL (2012: 1173). Nonetheless, Cuthona schraderi Eliot, 1907 View in CoL and Cuthonella paradoxa Eliot, 1907 . differs significantly from any species of the Since C. elioti View in CoL is known to have supplementary genus Guyvalvoria View in CoL by the presence of distinct gland that inserts into the penial base and ceratal rows and from G. “paradoxa ” sensu possibly a penial stylet, and C. modesta View in CoL is Valdés et al. (2012) also by considerably also described as having supplementary fewer lateral denticles ( Odhner, 1926).
gland that inserts into the penial base and Thus, Cuthona schraderi View in CoL is not a potential jaws without denticles (Eliot, 1907; Valdés synonym of G. francaisi Vayssière, 1906 View in CoL or et al., 2012), they are definitely not considered G. “paradoxa ” sensu Valdés et al. (2012: 1173).
to be Cuthonella . The taxonomic placement of According to the morphology of the radular these taxa should be addressed in a separate teeth and reproductive system, Guyvalvoria study. “paradoxa ” sensu Valdés et al. (2012) is
It has also subsequently been shown the most similar to G. gruzovi ( Martynov ,
that Cuthonella paradoxa belongs to a valid 2006a), but not to G. francaisi Vayssière, 1906 Antarctic genus Guyvalvoria Vayssière , (= “ Cuthonella” paradoxa Eliot, 1907 ).
1906, and has been synonymised with the The genus Guyvalvoria is a well-defined type species of the genus Guyvalvoria , G. taxon by the very specific combination francaisi Vayssière, 1906 because of the high of a posterior anus, laterally compressed morphological similarity ( Martynov , 2006a). branched rows of cerata, uniserial radula However, Guyvalvoria paradoxa sensu Valdés and a supplementary gland that inserts into et al. (2012: 1171–1173) does not match the the penial base. These features were already original description of Cuthonella paradoxa described in detail elsewhere ( Martynov ,
in Eliot (1907: 24–25) and considerably 2006a) and by those characters Guyvalvoria differs in the radular morphology. There is not only clearly different from the genus are seven distinct, “fairly stout and long” Cuthonella , but also from the majority of lateral denticles of the central teeth in the other aeolidacean families (Korshunova original description of Cuthonella paradoxa et al, 2017b; Martynov et al., 2020a). Some (Eliot 1907: 25, 11 mm preserved length) morphological similarity in presence of and 13–15 small denticles in Guyvalvoria the posterior anus Guyvalvoria shares only “paradoxa ” sensu Valdés et al. (2012: 1172, with the family Murmaniidae ( Martynov ,
15 mm preserved length). Notably, a fewer 2006a; Martynov et al., 2020a). Therefore, number (ca. 5–11) of distinct, long lateral Guyvalvoria Downloaded recently from was Brill.com transferred 12/12/2023 with 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ reservation to the family Murmaniidae type species apparently may also have a few (Korshunova et al., 2017b: Supplementary additional indistinct rows), an exclusively material1,Synopsisofthefamiliesofsuborder uniserial radula with a central cusp always Aeolidacea). The current status of the genus protruding (except for the few anteriormost Guyvalvoria as “taxon inquirendum” of eroded teeth), either with lateral denticles the “family Eubranchidae ” (MolluscaBase, closely adhering to the central cusp (as in 2020) should therefore be corrected to fully the type species) or usually distinct from valid status and, pending the availability of the central cusp. In the reproductive system, molecular data, to the family Murmaniidae . so far only a single receptaculum seminis The family Murmaniidae , on one hand, is reported (with a tendency to be more is morphologically consistent with the proximal in Cuthonella concinna s.l., otherwise characters of Guyvalvoria , but on the other in the majority of species it is distal) and the hand, is related to the families Cuthonellidae receptaculum seminis is usually oval, but it can and Xenocratenidae according to the most also be a rather tubular one (e.g., C. abyssicola recent phylogenetic analysis ( Martynov et s.l., C. punicea ). The penis is unarmed.
al., 2020a). Therefore a future phylogenetic The most specific feature of the genus – the analysis will show if Guyvalvoria can be insertion of the supplementary gland into the retained in the family Murmaniidae or vas deferens instead of the penis – was evident should be placed in its own family. in all the Cuthonella species that we were able
In the original diagnosis of the genus to examine (figs 4–20). Cuthona norvegica was Cuthonella, Bergh (1884) did not mention the originally described by Odhner (1929) within key feature that has proven to be present in the genus “ Cuthonella ”. However, C. norvegica all species of the genus Cuthonella studied so has a supplementary gland inserted into the far (except for uncertainties in the species C. penial base (Millen, 1986) and was transferred cocoachroma Williams & Gosliner, 1979 , see to the genus Cuthona s.str. by Martynov below), which is that the supplementary gland (1992). There are no new specimens of is inserted into the vas deferens instead of the Cuthona norvegica available, and after a future penis ( Martynov , 1992; present study). From clarification of its phylogenetic position, it the characters initially mentioned by Bergh may be necessary to transfer this species to a (1884), the dorso-lateral position of the anus different genus.
cannot be considered specific for the genus The insertion of the supplementary gland Cuthonella , and the absence of cnidosacs in into the vas deferens is likely a plesiomorphic the type species of the genus Cuthonella was feature because it is also present in at least disproved by Millen (1986). one very different taxon with a triserial radula So, at this time the genus Cuthonella should – Eubranchus tricolor – within the family be defined by the following features: relatively Eubranchidae (Korshunova et al., 2017a, b).
abundant anterior pre-anal rows of cerata, Potentially, this feature may occur within some either branched or unbranched (the majority other basal ‘tergipedids’. However, among of species have at least five to six, or more, genera and families with a uniserial radula it pre-anal ceratal rows in the adult state), a has so far been confirmed only for Cuthonella predominantly acleioproctic anus (the type and Cuthonellidae . Therefore, the insertion of species and a few other species may have a the supplementary gland into the vas deferens cleioproctic one), relatively broad jaws with should be used in combination with the other usually a single row of distinct denticles (the characters Downloaded mentioned from Brill above.com to12 identify /12/2023 future 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ members of Cuthonellidae . Notably, within the family Cuthonidae (genera Cuthona and Bohuslania ) this feature has so far never been observed (Korshunova et al., 2017a, 2018a).
The family Cuthonellidae Miller, 1977 has been resurrected recently, including a morphological diagnosis and molecular phylogenetic data (Korshunova et al., 2017a, b, 2018a). Miller (1977: 200) proposed the subfamily name Cuthonellinae based on the valid genus name Cuthonella and provided a brief morphological diagnosis “digestive ducts unbranched”. This diagnosis meets the requirements of the relevant ICZN (1999) articles (i.e., 11.7, 13, 29) for the availability of a family group name. Miller (1977: 200) also mentioned that the name Cuthonellinae “replaced” the name Precuthoninae introduced by Odhner in Franc (1968: 885). Miller did not provide explanations of his reasons and did not make a special indication why this is a “replacement” name. Bouchet & Rocroi (2005: 59) commented that Cuthonellinae Miller (1977) was invalid; they said in their remarks that it was “Introduced presumably (and thus in violation of Art. 40.1) as a replacement name for Precuthoninae, update to the knowledge of the genus because Cuthonella has precedence over Cuthonella since its description in 1884. All Precuthona Odhner, 1929 .” However, previously known and novel species are Precuthona Odhner, 1929 is not a synonym of presented below in alphabetical order (figs the genus Cuthonella Bergh, 1884 and therefore 4–18).
there is no violation of Article 40.1 of ICZN
(1999). The name Precuthona Odhner, 1929 Cuthonella abyssicola abyssicola Bergh ,
is a synonym of the genus Cuthona (Brown, 1884
1980), which belongs to the different family (figs 1, 2, 4, 19A, 21)
Cuthonidae , supported by morphological Cuthonella abyssicola Bergh, 1884: 24–26 , and molecular data (Korshunova pl. 10, figs. 1–3, pl. 11, fig. 2, pl. 12, figs. 9–
et al., 2017a, 2018a). The name Precuthoninae 13; Martynov , 1992: 23; Martynov , 2006a: Odhner in Franc (1968: 885), therefore, is a 73; Martynov , 2006b: 290; Martynov & synonym of the family Cuthonidae Odhner, Korshunova, 2011: 195 .
1934. The name Cuthonellinae Miller, 1977 is Cuthonella berghi Friele, 1903: 11 , tab. II, fig.
thus an available family group name in full 3–4, tab. III, fig. 7–10.
compliance with Article 13 (“to be available, Cuthonella ferruginea Friele, 1903: 10 , tab. II, every new family-group name published after fig. 1–2, tab. III, fig. 3–5.
1930 must satisfy the provisions of Article Cuthona abyssicola (Bergh, 1884) – Millen, 13.1 and must be formed from an available 1986: 1358–1359, figs 8, 9.
genus-group name then used as valid by Type material. Holotype lost (K. Jensen the author in the family-group taxon [Arts. ( ZMUC), personal communication; K. Lundin 11.7.1.1, 29]”, ICZN 1999 ). The available family performed an additional search in ZMUC and group name Cuthonellinae Miller, 1977 (in a confirmed the absence of the type material).
restricted sense compared to the mention in Material. ZMMU Op-616, L = 23 mm Miller (1977), without “ Precuthona ”) with an length (preserved, convoluted, more than amended diagnosis was therefore introduced 30 mm if unrolled), Arctic Ocean , Barents as the valid family Cuthonellidae Miller, 1977 Sea , 77° 06´N, 58° 12´E, 307 m, 07.08.1948 GoogleMaps .
and the (incorrect and unnecessary) term NTNU-VM 73124 View Materials , 1 spec., L = 31 mm length “replaced” should be discounted. (preserved), Norwegian Sea, Nyegga, 64° Cuthonellidae is distinct by the 40.115N 5° 15.702E, 731 m, coll. Hans Tore morphological and molecular phylogenetic Rapp, 05.08.2008 GoogleMaps .
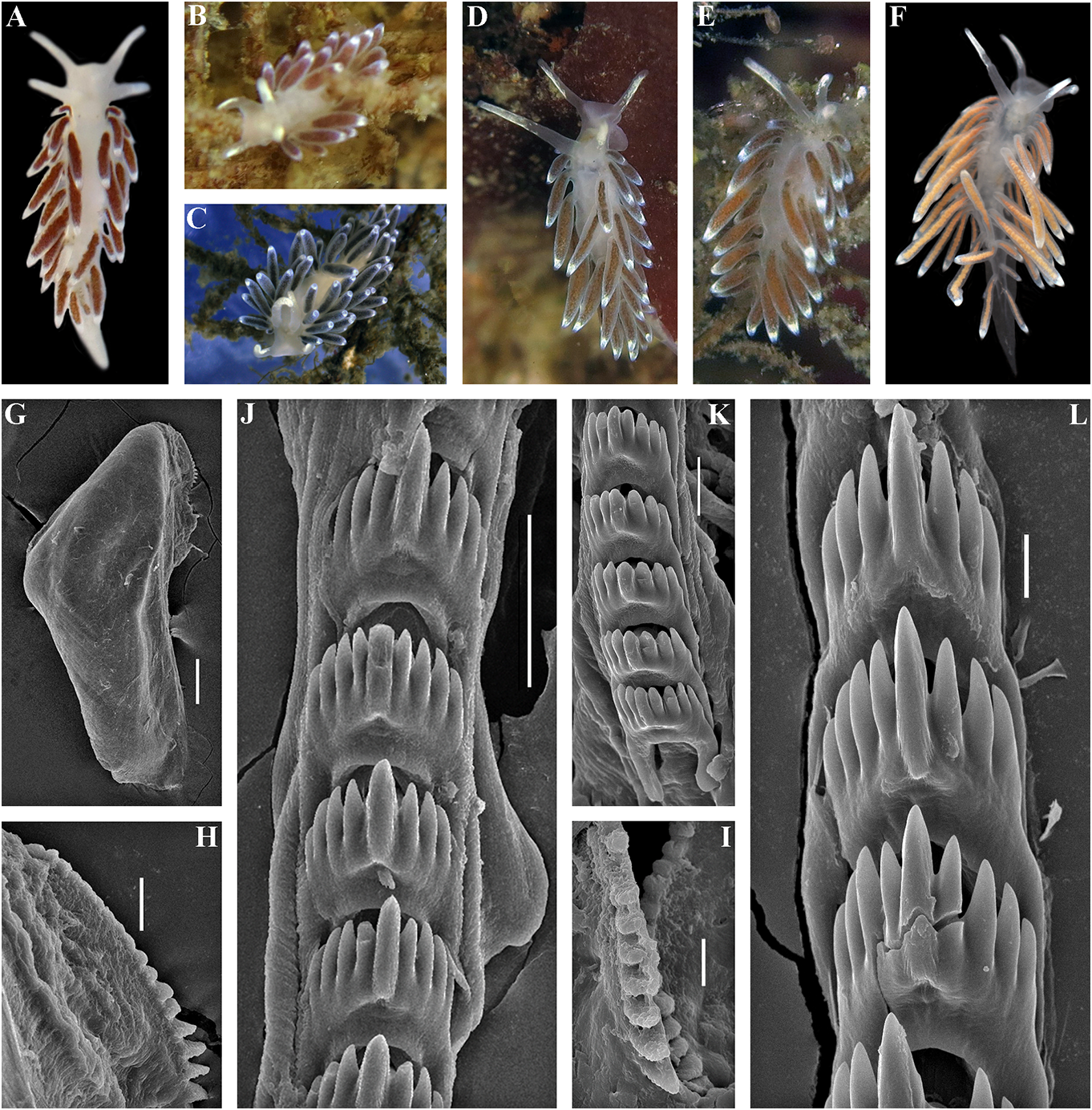
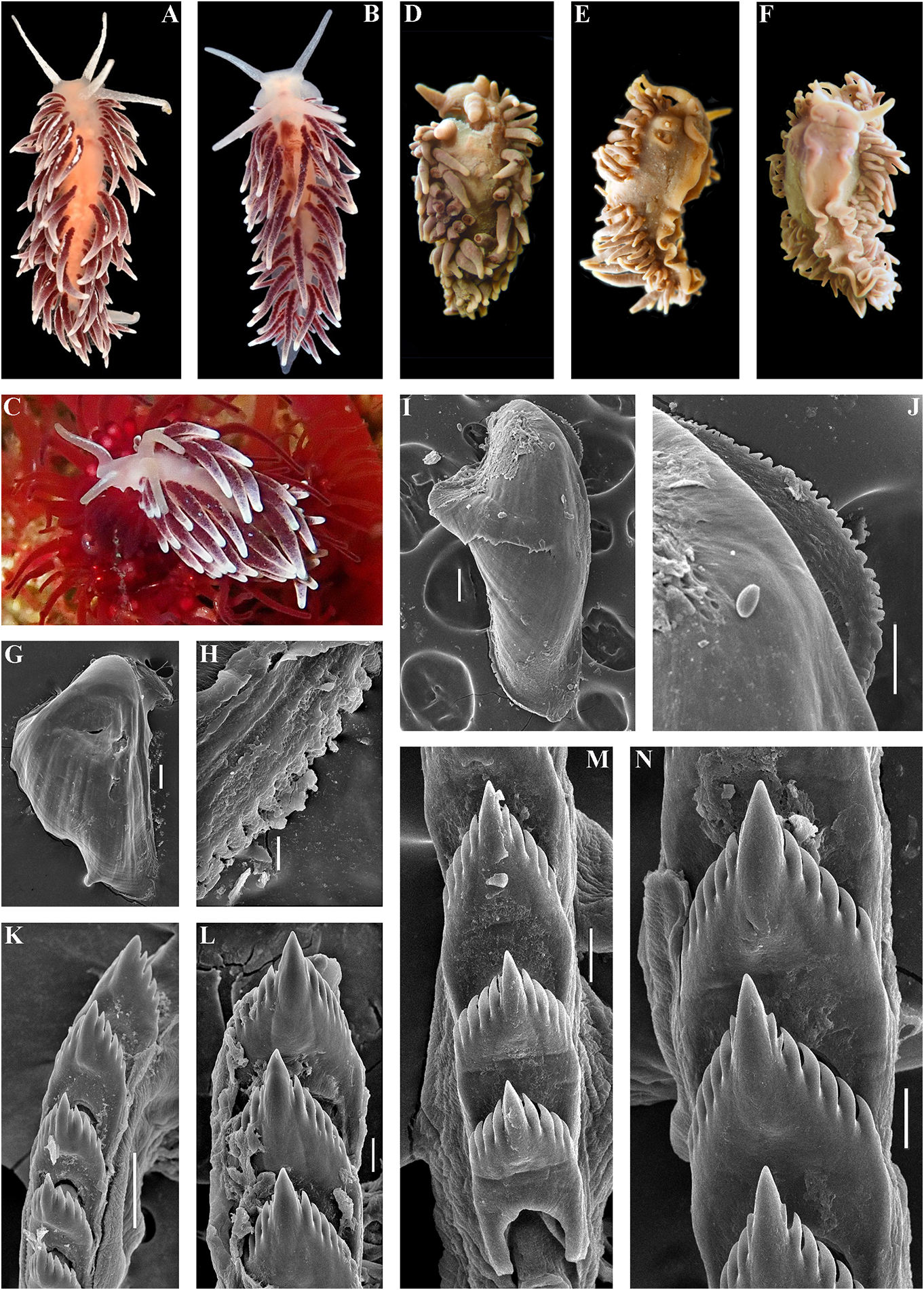
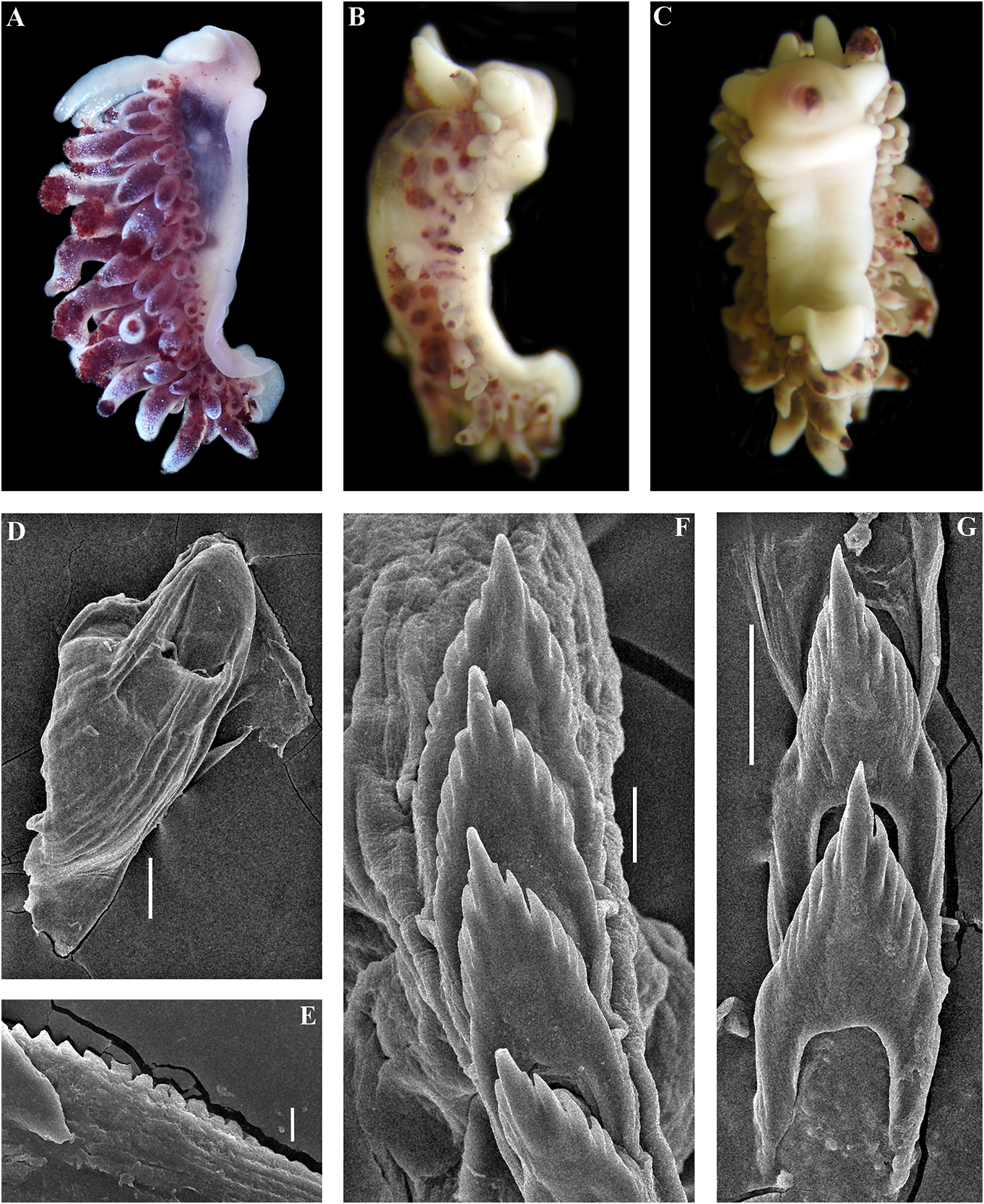
data (Korshunova et al., 2017a, 2018a, this External morphology. Body wide, massive.
study). In some publications (Korshunova et Foot and tail broad. Oral tentacles moderate.
al., 2017a, b) the year of the original publication Rhinophores smooth to slightly wrinkled,
of Cuthonellinae was indicated as ‘Miller, similar in size to oral tentacles. Dorsal cerata 1971’ due to a typo (the reference ‘Miller, 1977’ elongate, thick, arranged in continuous rows.
was provided correctly). The correct year of Up to 17 pre-anal branched ceratal rows. Anal publication is Miller (1977). The type genus of opening cleioproctic .
the family Cuthonellidae is Cuthonella Bergh, Colour (preserved). Background colour 1884. yellowish white. Digestive gland diverticula
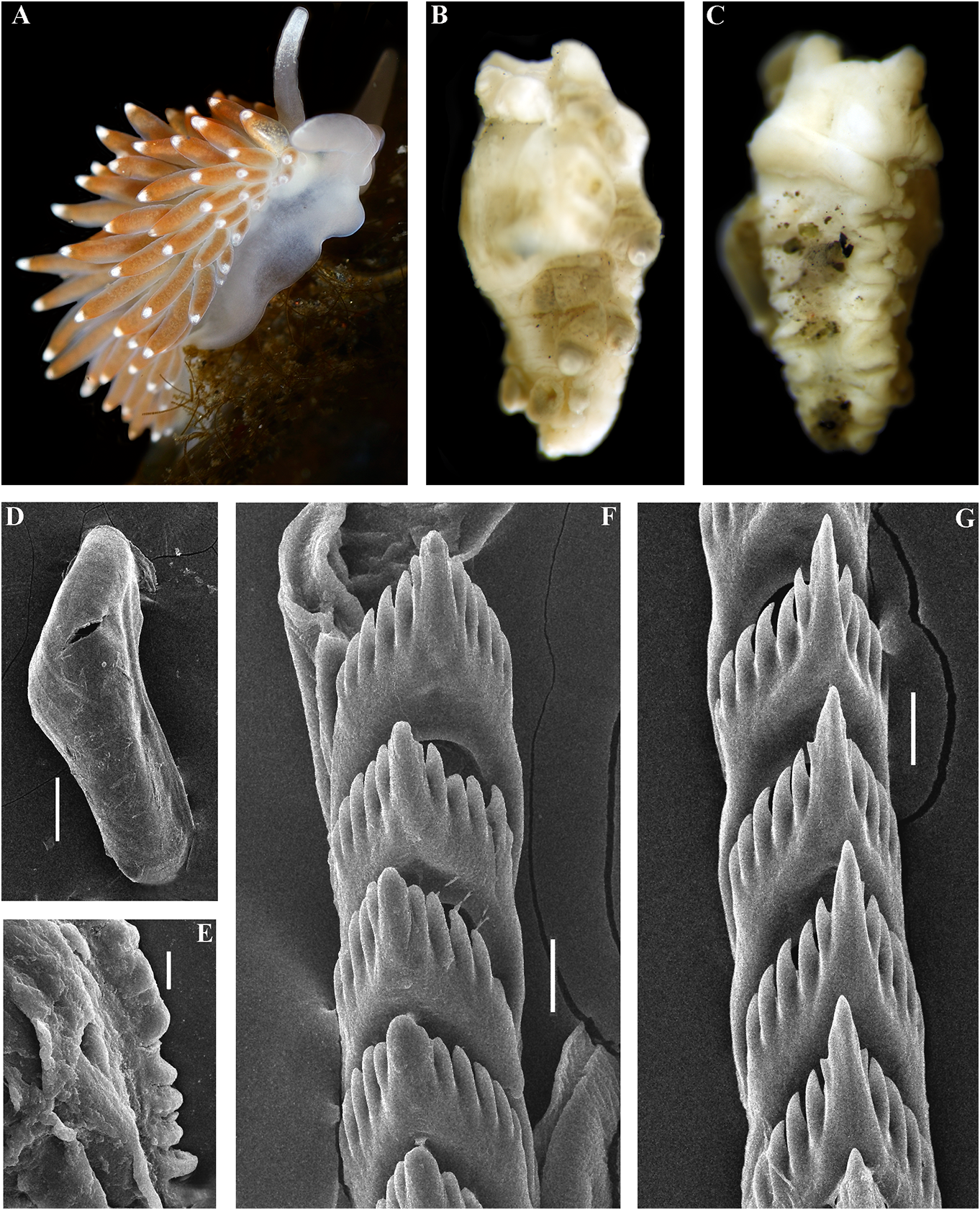
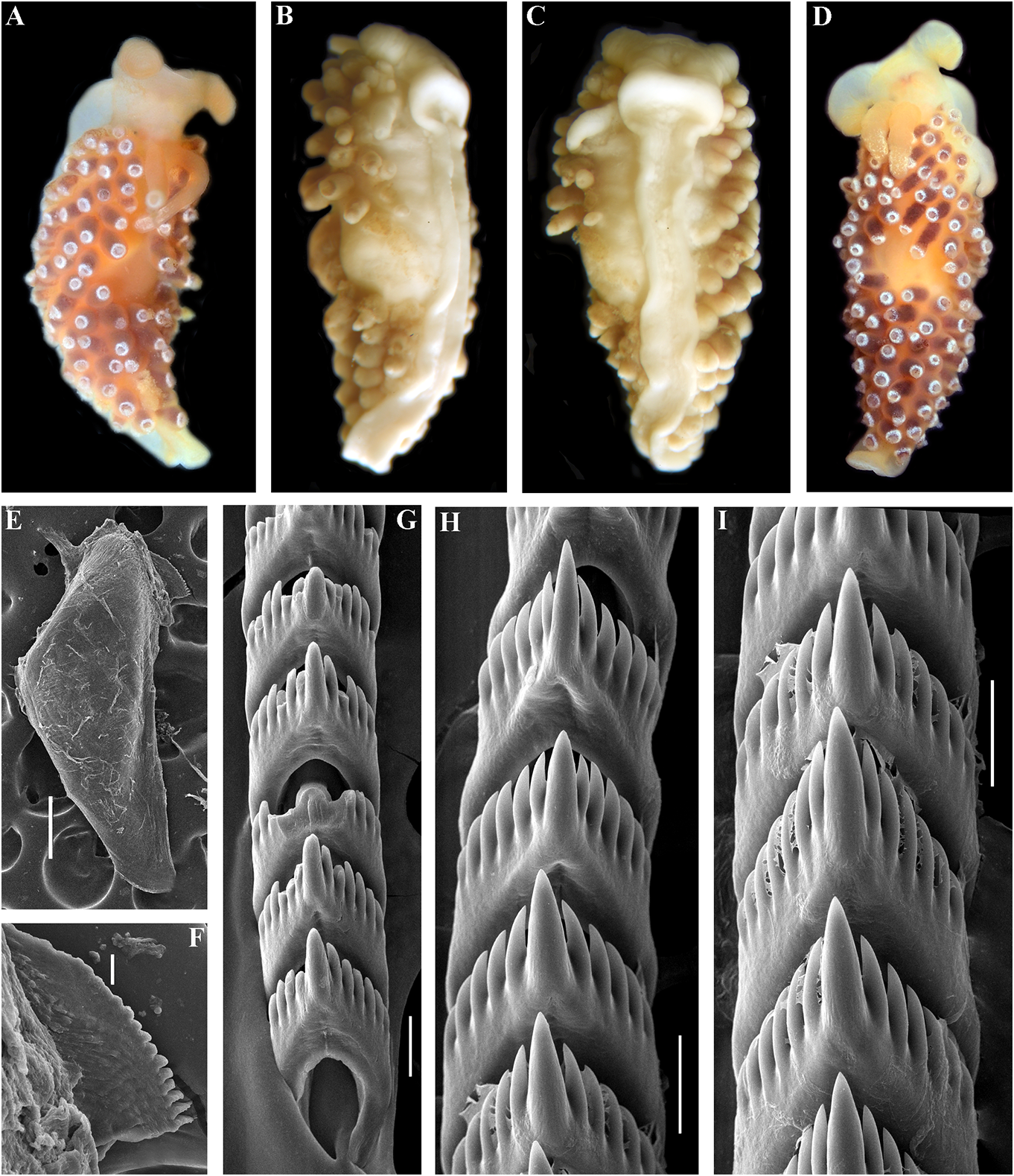
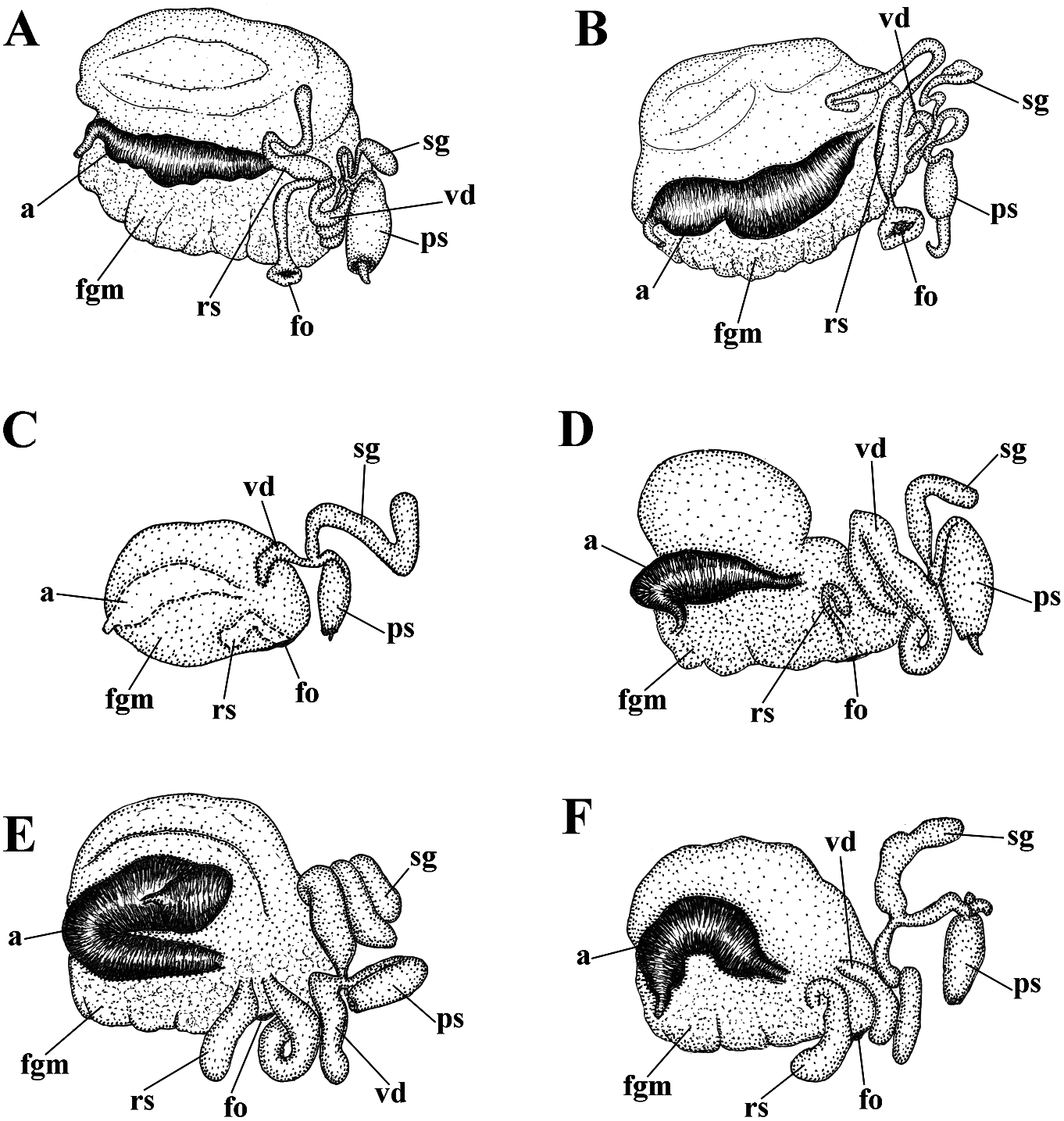
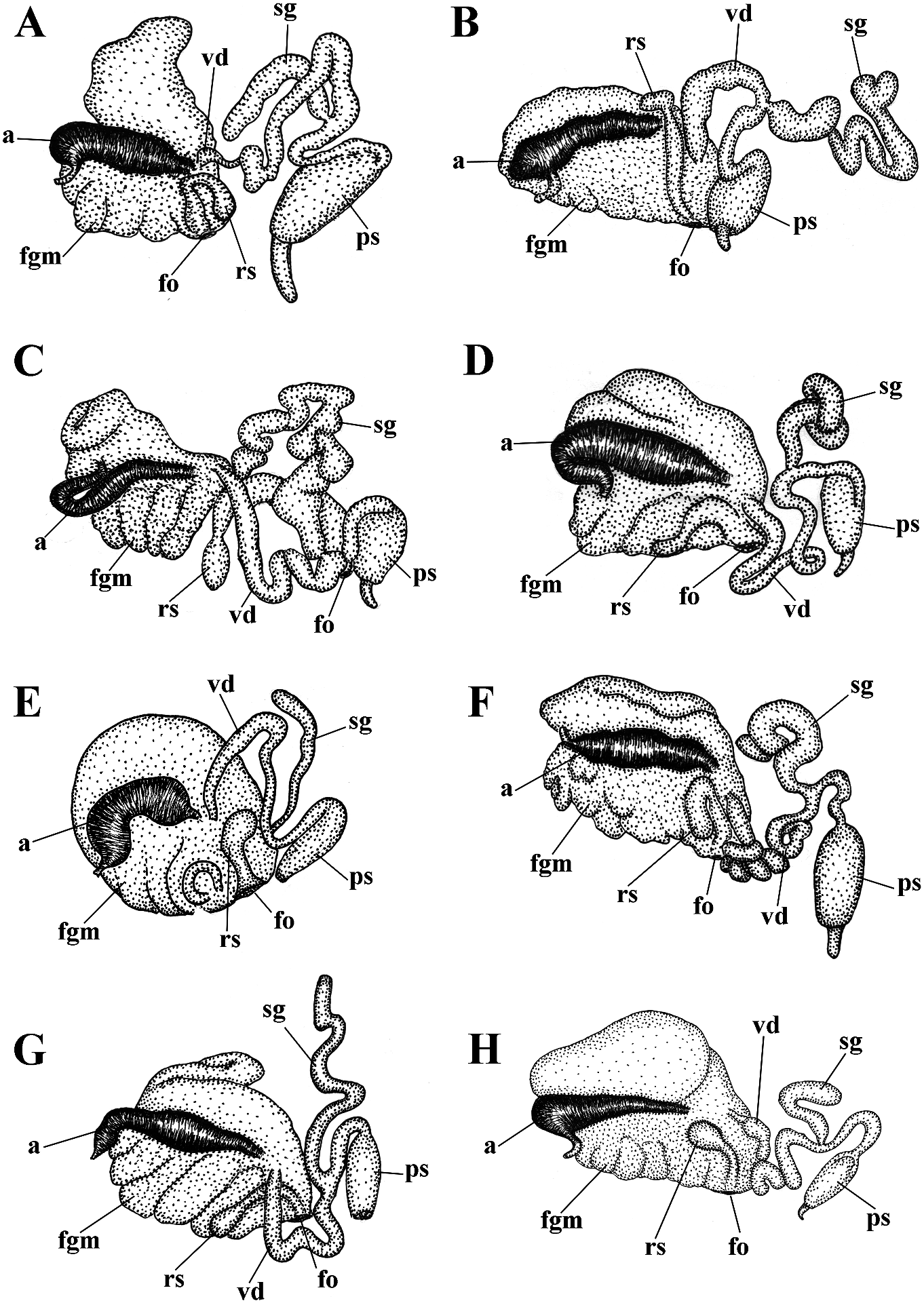
In the present revision of the genus in cerata possibly reddish to pinkish. Ceratal Cuthonella the number of known species is tips without distinct spots or bands of opaque increased threefold. It is the most substantial white pigment Downloaded. from Brill.com 12/12/2023 04:16:22PM via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/ Jaws. Jaws broad, yellowish to brown in specimen showed closely similar features colour. Masticatory processes of jaws covered with the deeper-water specimens (300 m) with several rows of denticles (with a few from the Barents Sea in the presence of indistinct additional rows) (fig. 4C–E). up to 17 pre-anal branched ceratal rows, Radula. Radula formula 21 × 0.1.0(specimen a long vas deferens and a receptaculum 23 mm in length, Barents Sea). Central tooth seminis on a long stalk. In the original broad, with a strongly protracted, pointed description (Bergh, 1884), a type specimen non-compressed cusp (fig. 4F, G). Central from deeper waters of the Faroe Channel teeth with up to 14 lateral denticles, which are (c. 1000 m) shows closely similar external rarely clustered. Cusp quite clearly delineated and reproductive features with the shallowfrom the adjacent lateral denticles which do water specimen from Franz Joseph Land, but not reach the top of the cusp. the radular teeth have a significantly fewer Reproductivesystem. Diaulic.Hermaphroditic number of lateral denticles (10–14 vs. 13–36). duct leads to a swollen ampulla. Vas deferens Notably, in the Franz Joseph Land specimen long, withoutadistinctprostate.Supplementary the lateral denticles are usually clustered gland long, inserts into the vas deferens at a and extend to the top of the central cusp considerable distance from the elongated penial (fig. 5I, J), compared to the deep-sea C. sheath. Penis conical, strong, unarmed. Oviduct abyssicola (fig. 4F, G). Millen (1986) performed connects through the insemination duct into a detailed investigation of the surviving type the female gland complex. Exceedingly long material of the nominal species Cuthonella tubular receptaculum seminis placed distally, berghi Friele, 1903 and C. ferruginea widened in the middle part with a small oval Friele, 1903, which have been considered reservoir (fig. 19A). synonymous with C. abyssicola . Both of these Ecology. Lives on soft bottoms with stones, synonymous species come from similar commonly at bathyal and shelf depths of c. geographic regions (NE Atlantic, Norwegian 300–1112 m. A record from the shallow waters Sea) and also from similar depths (from of the Laptev Sea ( Martynov & Korshunova, deeper waters of 590–1098 m) as C. abyssicola . 2011) (49 m depth) needs verification. The radulae of C. berghi and C. ferruginea Distribution. Faroe Islands in the North are similar to the original description of C. Atlantic; Norwegian Sea; Barents Sea, possibly abyssicola (Bergh 1884) : the central teeth the Laptev Sea (Millen, 1986; Martynov & possess only a few lateral denticles (c. 6–10), Korshunova, 2011; present study). and the cusp is more clearly delineated. This Remarks. Cuthonella abyssicola differs from agrees well with our specimens from the deep the majority of other Cuthonella species by the waters of the Barents Sea (c. 6–14) (fig. 4F, substantial number of anterior ceratal rows G). The number of teeth of the radula of the (up to 17), radular teeth with small denticles deeper water specimen from the Barents Sea which are not always clearly delineated from (21 teeth) also corresponds well with the first the cusp, long vas deferens and distal seminal description of Bergh (1884) and also to those receptacle on a very long stalk. described by Friele (1903) and Grieg (1913) In the present study, we give for the first (18–21 teeth), but not to the shallow-water time both morphological and molecular specimen from Franz Joseph Land (27 teeth). data for a relatively shallow-water specimen Thus, the radular characters (which are from Franz Joseph Land in the Arctic (see reliably based on several deep-sea specimens description below). Morphologically, this collected in Downloaded various from locations Brill.com) are different 12/12/2023 04:16:22PM
via Open Access. This is an open access article distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/
from the shallow-water specimen from Franz nudibranch taxa, such as in the genera Joseph Land, which by other features are Adalaria , Microchlamylla , and Ziminella . We similar to the real C. abyssicola . Therefore, have also studied a single specimen from the these differences cannot be ignored while Laptev Sea, from a relatively shallow-water making a taxonomic decision. Because only a locality at c. 50 m. Remarkably, compared to single shallow-water specimen was available, the specimen from Franz Joseph Land, the we preferred in this case to separate it here Laptev Sea specimen possesses no more than as a subspecies; Cuthonella abyssicola kryos 14–15 lateral denticles on the central tooth subsp. nov. (see description below). (23 teeth in total), and thus more closely
It was previously shown (Korshunova et matches the real deep-sea C. abyssicola al., 2017b; Martynov & Korshunova, 2017) that than the Franz Joseph Land specimen. Thus, the waters of Franz Josef Land demonstrate the distributional Downloaded from data Brill.com for 12 C /12. /2023 abyssicola 04:16:22PM considerable endemism in other via Open various Access. This abyssicola is an open and access C. abyssicola article distributed kryos subsp under.the nov terms. of the CC-BY 4.0 license. https://creativecommons.org/licenses/by/4.0/
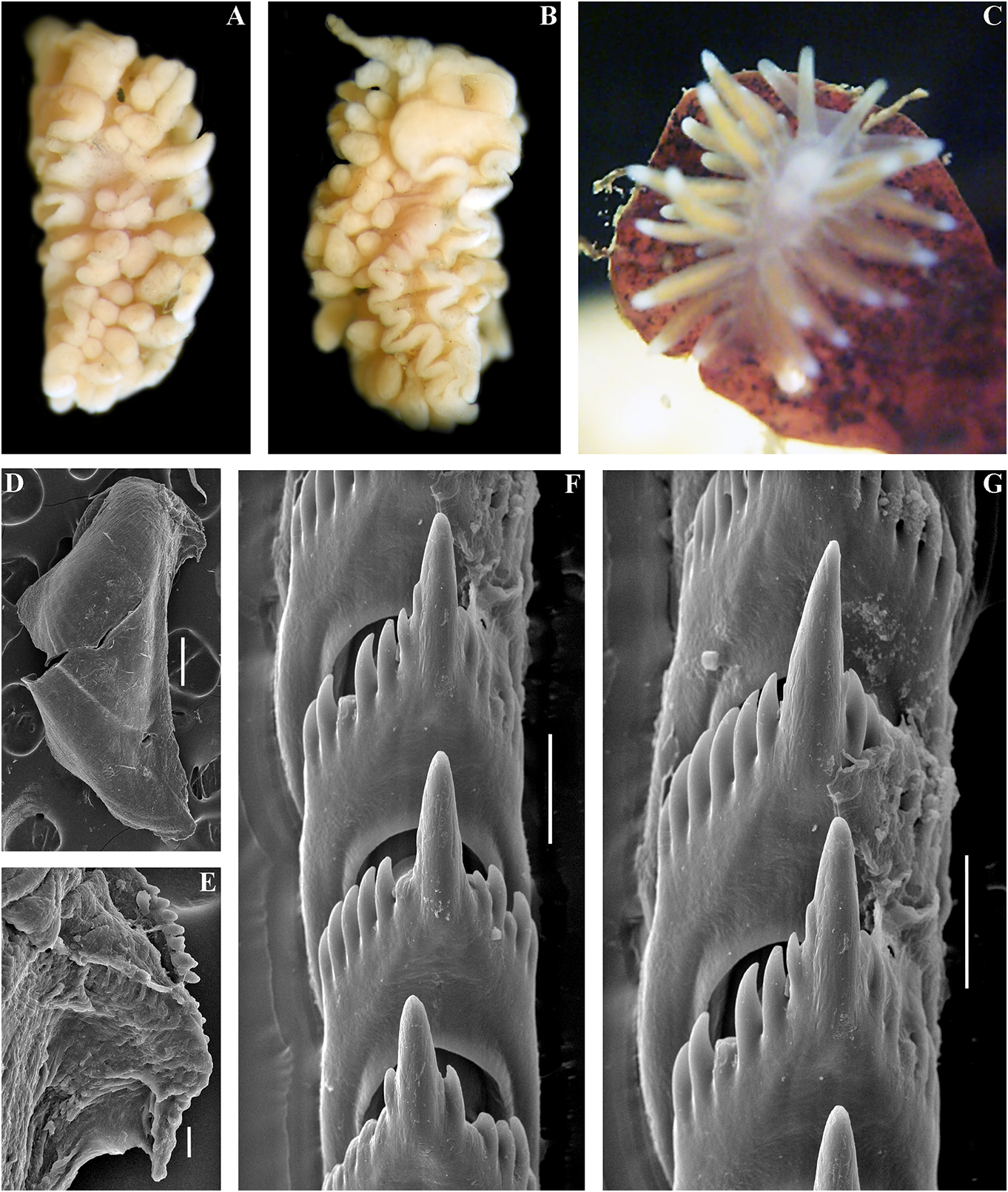
implies that the nominative subspecies can smooth to slightly wrinkled. Dorsal cerata be found in shallow depths, but not in the elongate, thick, arranged in continuous rows. high latitude Franz Joseph Land. It should Up to 17 pre-anal branched ceratal rows. Anal also be taken into consideration that the opening cleioproctic. shallow-water record from the Laptev Sea Colour (live). Background colour light needs further verification. C. abyssicola s.l. pinkish to whitish. Digestive gland diverticula potentially feeds on athecate hydroids. We in cerata light pinkish to whitish. Ceratal tips have studied few deep-sea specimens of with dispersed white pigment. Rhinophores C. abyssicola abyssicola , but none of them light yellowish with indistinct white pigment were suitable for molecular investigation towards the top (fig. 5A). due to the considerable age of collection Jaws. Jaws broad, yellowish to brown in and primary fixation in formaldehyde. colour. Masticatory processes of jaws covered We made additional inquiries to obtain with a few rows of denticles (with a few recent deep-sea specimens, but none were indistinct additional rows) (fig. 5F, G, H). available. Therefore, though details of the Radula. Radula formula 27 × 0.1.0. Central radular features and distributional patterns tooth broad, with a strongly protracted, may potentially imply that we are dealing pointed non-compressed cusp (fig. 5I, J). with two sister species rather than with two Central teeth with up to 36 lateral denticles subspecies, we need more data to confirm (often clustered and increasingly irregular this. However, because the shallow-water towards the cusps). Cusp is not clearly subspecies (see below) shows substantially delineated from the adjacent lateral denticles similar major features in both external which may reach the top of the cusp. and internal morphology, including the Reproductive system. Diaulic. reproductive system, we can confirm that it Hermaphroditic duct leads to a long, represents a form that is close to the original moderately swollen convoluted ampulla. Vas Cuthonella abyssicola and therefore can be deferens long, without a distinct prostate. used here for defining the genus Cuthonella . Supplementary gland long, inserts into the vas For a morphological comparison with other a considerable distance from the elongated Cuthonella species, see table 2. penial sheath. Penial sheath elongated. Penis conical, strong, unarmed. Oviduct connects Cuthonella abyssicola kryos subsp. nov. through the insemination duct into the
( Figs 1 View FIGURE 1 , 2 View FIGURE 2 , 5 View FIGURE 5 , 19B View FIGURE 19 , 21 View FIGURE 21 ) female gland complex. Very long tubular ZooBank: http://urn:lsid:zoobank. receptaculum seminis in a distal position, o r g: a c t:9 F 7 FA 1 E 7- 3 3 2 6 - 4 0 0 9- 9 F 9 B - with a widened base and small pear-shaped ADFFC10E7F82 reservoir (fig. 19B). Type material. Holotype, ZMMU Op-727, Ecology. Lives on soft bottom with stones at L = 25 mm length (preserved), Arctic Ocean, depths of 18– 33 m. Franz Josef Land, Wiltona Island, 18–33 m Distribution. High latitude arctic region of depth, collected by O.V. Savinkin, 23.08.2013. Franz Josef Land. Etymology. From Ancient Greek ΚΡΎΟç Remarks. See remarks for C. abyssicola (kryos), “icy cold” in reference to the Arctic, abyssicola .The lowest COI intergroup distance high latitude type locality. of 7.00% is found between C. abyssicola kryos External morphology. Body wide, massive. subsp. nov. and C. sandrae sp. nov. (table 1). Foot and tail broad. Oral tentacles moderate. For a morphological Downloaded from comparison Brill.com with 12/12/2023 other 04:16:22PM Rhinophores similar in size to oral via tentacles Open, Cuthonella Access. This is an open species access, see article table 2. distributed under the terms of the CC-BY 4.0 license.
https://creativecommons.org/licenses/by/4.0/




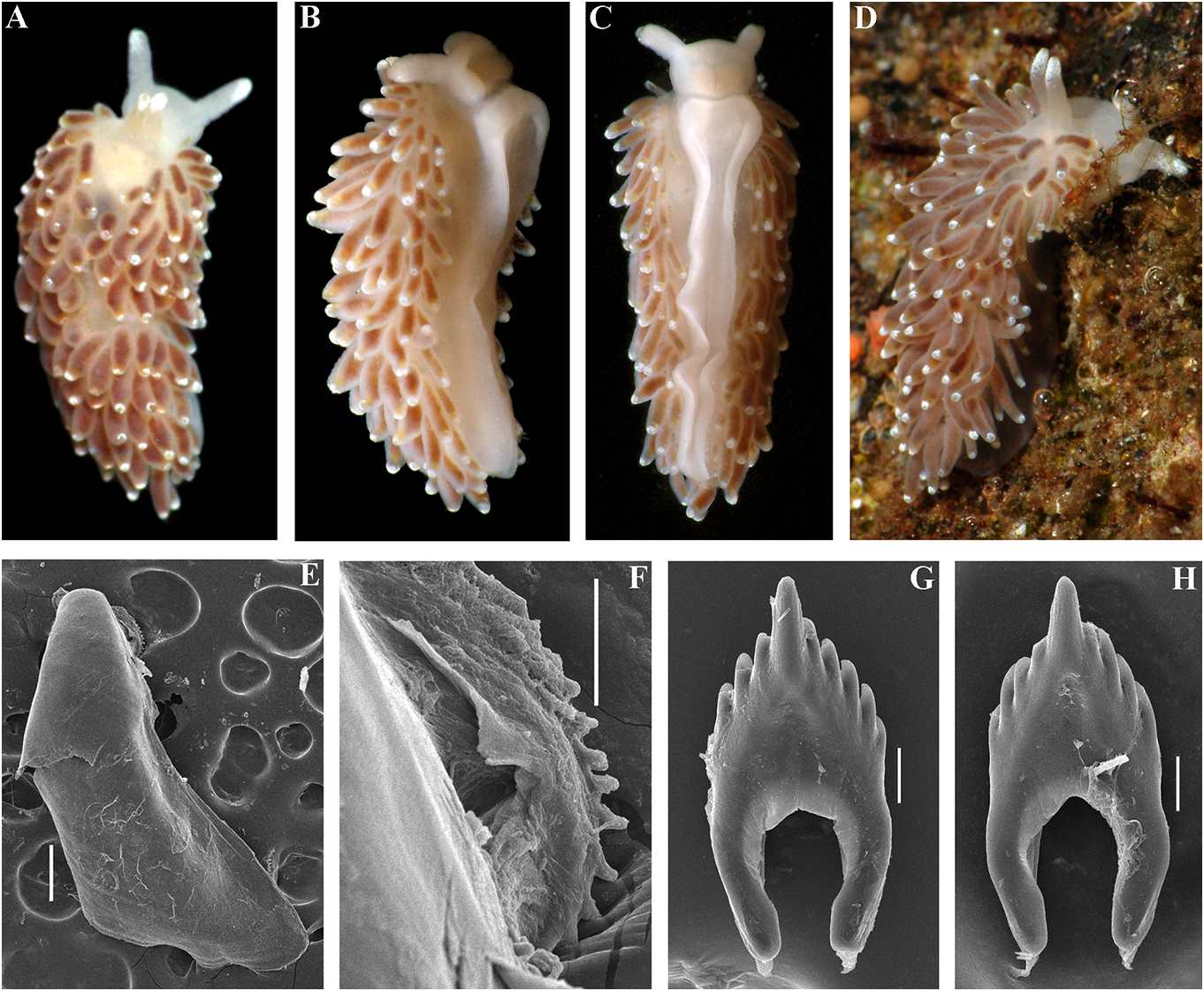





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cuthonella concinna
| Korshunova, Tatiana A., Sanamyan, Nadezhda P., Sanamyan, Karen E., Bakken, Torkild, Lundin, Kennet, Fletcher, Karin & Martynov, Alexander V. 2021 |
Guyvalvoria gruzovi
| Martynov 2006 |
Cuthonella
| Martynov 1992 |
C.soboli
| Martynov 1992 |
C. soboli
| Martynov 1992 |
Cuthonella
| Martynov 1992 |
Cuthona
| sensu Millen 1986 |
Cuthonella modesta
| Eliot 1907 |
Cuthona schraderi
| Eliot 1907 |
Cuthonella paradoxa
| Eliot 1907 |
G. “paradoxa
| Eliot 1907 |
C. modesta
| Eliot 1907 |
Cuthona schraderi
| Eliot 1907 |
G. “paradoxa
| Eliot 1907: 1173 |
G. francaisi Vayssière, 1906
| Vayssiere 1906 |
G. francaisi Vayssière, 1906
| Vayssiere 1906 |
Cuthonella abyssicola
| Bergh 1884 |