
Emersoniella reninoda Gustafsson & Bush
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3796.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:D3E1601F-74E9-4594-8820-F5365DB42AF1 |
|
DOI |
https://doi.org/10.5281/zenodo.6127089 |
|
persistent identifier |
https://treatment.plazi.org/id/03EF879B-8249-2540-5CFF-4E6D1E7D79D2 |
|
treatment provided by |
Plazi |
|
scientific name |
Emersoniella reninoda Gustafsson & Bush |
| status |
sp. nov. |
Emersoniella reninoda Gustafsson & Bush , new species
( Figs 1 View FIGURE 1 , 3 View FIGURE 3 a–d)
Type host. Melidora macrorrhina macrorrhina Lesson, 1827 ( Coraciiformes : Alcedinidae ) – hook-billed kingfisher.
Diagnosis. Emersoniella reninoda new species keys out to couplet number 3 ( E. halcyonis and E. regis ) in the key of Emerson & Price (1978) but, based on the abdominal chaetotaxy, it is more similar to E. halcyonis than to E. regis , although it has even more setae on tergites VI–VII than those two species. Similar to E. crassicarina , but differs in abdominal chaetotaxy and the shape of the male genitalia, with the penis of E. crassicarina not reaching the endomeral plate, as is the case in E. reninoda . Further, E. crassicarina lacks the dorsal anterior suture of E. reninoda .
Description. Head roughly triangular, with rounded temples and broadly flattened frons ( Figs 1 View FIGURE 1 , 3 View FIGURE 3 a). Marginal carina interrupted laterally and submedianly at feeding canal, and with several small carinal islands median to marginal carina. Posterior to lateral interruption marginal carina is broad, with irregular median margin. Premarginal carina small, triangular. At feeding canal, marginal carina is displaced deeply posteriorly and dorsally, and widened into broad arch. Frons broadly hyaline, continuous laterally with dorsal preantennal suture that arises from lateral interruptions of marginal carina, and then continues posteriorly to site of ADS, which they envelope. Sutures do not connect medianly, and dorsal anterior plate is thus not cut off from main head plate. AS 2-3 on premarginal carina, AS 1 and DSMS near anterior end of postmarginal carina, each with attendant canals in marginal carina. VSMS1-2 equidistant from anterior margin of head. Ventral carinae convergent to anterior margin of pulvinus, where they turn and run in parallel, but are diffuse and not clearly connected to marginal carina anteriorly. AVS3 anterior to AS 1, AVS1-2 positioned on ventral side of marginal carina, each with attendant canals in carina. Preantennal nodi large, kidney-shaped. Coni shorter than scapes, roundedly triangular ( Fig. 1 View FIGURE 1 ). PNS and one pair of PTS relatively long. Preocular nodi elongated, not very distinct. Postocular nodi not distinct from marginal temporal carina, which is narrow and continuous around post-antennal head. POS and OS both located on eye, OS long, POS spine-like. MTS1 long setae, perhaps two thirds of the length of MTS3; other MTS short and slender. Gular plate slender, spade-shaped, with concave antero-lateral margins.
Prothorax shorter than wide, with one pair of short setae on posterior margin, median to spiracle openings. Prosternum nude. Proepimera broad, with blunt median ends. Pterothorax wider than long, with divergent lateral margins and posterior margin convergent to median point. Mesosternum with two pairs of short setae. Metasternum large, more or less circular, with five pairs of short setae distributed around lateral and posterior margins ( Fig. 3 View FIGURE 3 b). Tergites II–VIII divided medianly in both sexes ( Fig. 3 View FIGURE 3 b).
Male. Posterior margin of pterothorax with 10–11 pairs of setae. Pleurites III–V with well-developed, hookshaped re-entrant heads, pleurites VI–VIII with well-developed, flat heads that are not, or only barely, re-entrant. Abdominal chaetotaxy as in Table 3 View TABLE 3 . Male genitalia ( Fig. 3 View FIGURE 3 c): basal plate long, beginning in middle of segment VI, but diffuse anteriorly. Lateral margins thickened in distal half; these thickenings are continuous with lateral and median thickenings of parameres, which are fused with basal plate. Medianly basal plate open to an oval atrium, in which the mesomere and endomere are located. Endomere broader than long, with antero-lateral extensions that reach to site of parameral fusion. One pair of distinct nodi in postero-lateral corner of endomere, connected medianly by a rugose thickening. Mesomere-proper as four needle-like bars, the two lateral pairs about a fourth of the length of the median pair; does not reach or just barely reaches anterior margin of endomere. Antero-lateral sclerites broad, somewhat club-shaped, and slightly thickened anteriorly. One pair of minute setae near distal tip of parameres, and one pair of small pores lateral to this, both on outer margin. Measurements (n = 15): TL: 1.75–1.92 (1.84); HL: 0.54–0.59 (0.56); HW: 0.57–0.61 (0.59); PRW: 0.31–0.33 (0.32); PTW: 0.45–0.50 (0.47); AW: 0.61–0.67 (0.65).
Female. Posterior margin of pterothorax with 10 pairs of setae. Pleurites III–VI with well-developed, hookshaped re-entrant heads, pleurites VII–VIII with well-developed, flat heads that are not, or only barely, re-entrant. Abdominal chaetotaxy as in Table 4 View TABLE 4 . Vulval margin with 6–8 marginal pairs of long, slender setae, and seven submarginal pairs of shorter setae, which are not thorn-like. Oblique set with three pairs of slender setae, situated far from vulval margin. Three pairs of long, stout setae postero-lateral to vulval margin ( Fig. 3 View FIGURE 3 d). Measurements (n = 15): TL: 1.92–2.17 (2.07); HL: 0.57–0.62 (0.59); HW: 0.062–0.66 (0.64); PRW: 0.33–0.36 (0.35); PTW: 0.49–0.54 (0.52); AW: 0.67–0.80 (0.72).
Type material. Male holotype, ex Melidora macrorrhina macrorrhina , PAPUA NEW GUINEA: Northern District, Popondetta, Jumbora Plantation, elev. 200 ft, 25. IX. 1963, H. Clissold, host BBM-NG-29668 ( NHML). Female allotype, on same slide as holotype ( NHML). Paratypes: 2 males, same data as holotype (both slides also contain female E. crassicarina ) ( NHML). 3 males and 3 females, same data as holotype ( USNM). 3 males and 1 female, same data as holotype (PIPeR).
Non-type material. 1 nymph, on same slide as holotype. 2 nymphs, same data as holtype PAPUA NEW GUINEA: Morobe District: 15 males, 25 females, 4 nymphs, 10 km W of Bulolo, elev. 780 m, 13. VIII. 1967, A.B. Mirza, host BBM-NG-54237 ( NHML). PAPUA NEW GUINEA: Northern District: 4 males, 4 females, 1 nymph, Cape Killerton, elev. 10 ft, 22. X. 1963, H. Clissold, host BBM-NG-29287 ( USNM). 2 males and 6 females, Popondetta, Jumbora District, elev. 25 m, 2. VI. 1966, G. Lippert, host BBM-NG-25017. 23 males, 46 females, 7 nymphs, Popondetta, Jumbora Plantation, 5. V. 1966, P.J. Shanahan, host BBM-NG-24885. 2 females, Vicinity of Popondetta, Empi Lakes, elev. 300 ft, 24. X. 1963, H. Clissold, host BBM-NG-29321; two unidentified nymphs belonging to the “ Degeeriella complex” on same slide. PAPUA NEW GUINEA: Western District: 1 nymph, Oriomo River, elev. 20 ft, 14. II. 1964, H. Clissold, host BBM-NG-29520 (all at PIPeR).
Etymology. The species epithet is formed by “ ren ”, Latin for “kidney”, and “ nodus ” for Latin “knot”, referring to the kidney-shaped preantennal nodi.
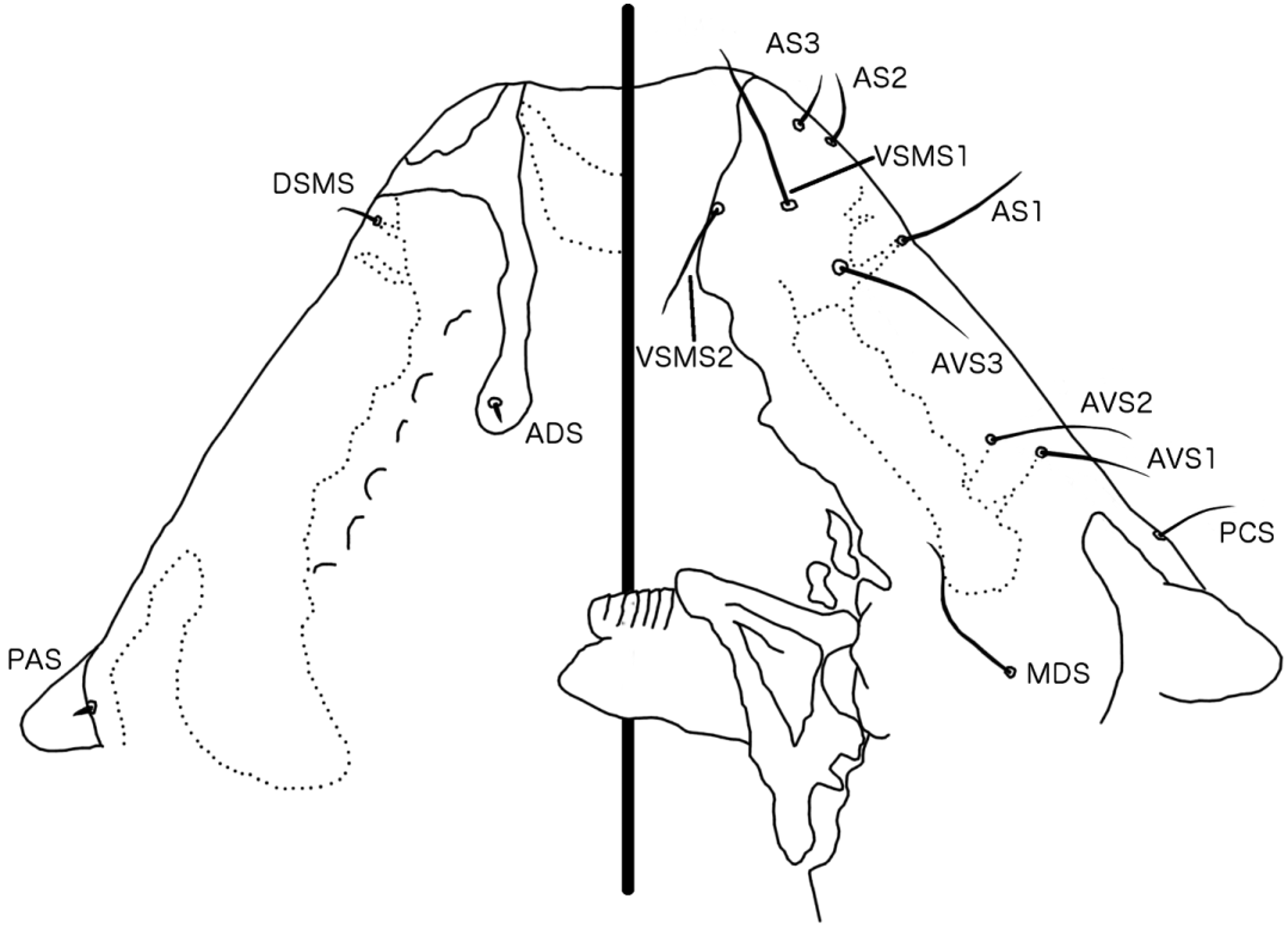
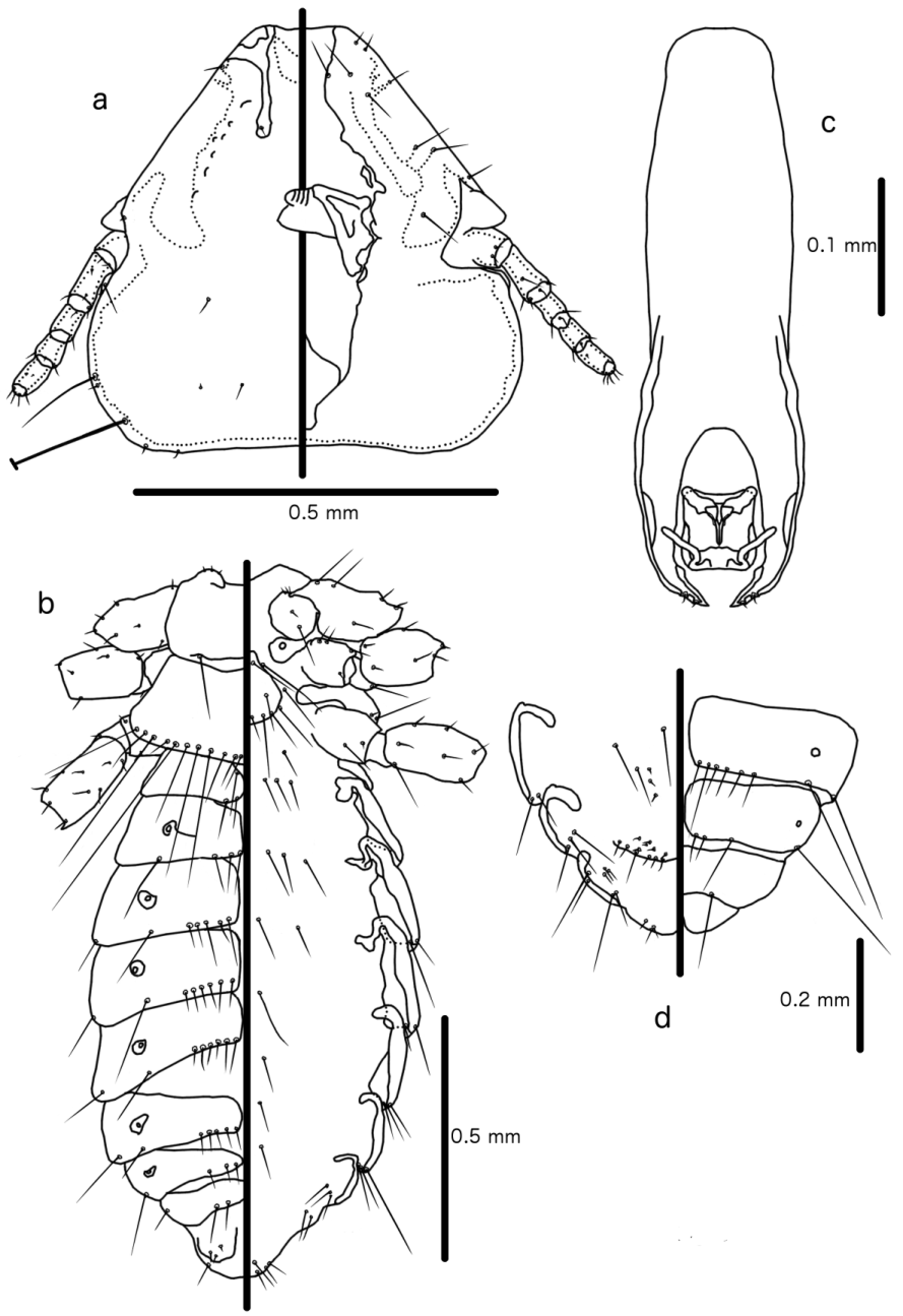
TABLE 3. Male abdominal chaetotaxy for Emersoniella reninoda n. sp. (see explanatory notes in Table 1).
| Segment | Tergomedian | Post-spiracular | Sternal | Postero-lateral |
|---|---|---|---|---|
| II | 1 + 2 | 0 | 1 + 3 | 0 |
| III | 3 | 1 | 3 | 0 |
| IV | 5 | 1 | 2 | 3 |
| V | 6 | 1 | 1 | 3 |
| VI | 6 | 1 | 1 | 4 |
| VII | 5 | 1 | 1 | 4 |
| VIII | 3 | 0 | 1 | 4 |
| IX | 2 | 0 | 0 | 3 |
| Terminal | 0 | 0 | 2 | 3 |
TABLE 4. Female abdominal chaetotaxy of Emersoniella reninoda n. sp. (see explanatory notes in Table 1).
| Segment | Tergomedian | Post-spiracular | Sternal | Postero-lateral |
|---|---|---|---|---|
| II | 1 + 2–3 | 0 | 1+3 | 0 |
| III | 3–4 | 1 | 3 | 0 |
| IV | 4–5 | 1 | 2 | 3 |
| V | 4–5 (6) | 1 | 1 | 4 |
| VI | 5 | 1 | 1 | 4 |
| VII | 5 (6–7) | 1 | 3 | 4 |
| VIII | 3 | 0 | 0 | 4 |
| IX | 1 | 0 | 0 | 2+2 |
| Terminal | 0 | 0 | 3 | 0 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.