
Fortiblatta cuspicolor Liang, Vršanský et Ren, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.185099 |
|
DOI |
https://doi.org/10.5281/zenodo.6225514 |
|
persistent identifier |
https://treatment.plazi.org/id/E60787D0-FF9D-FFBD-FF11-74D4ADC91FF5 |
|
treatment provided by |
Plazi |
|
scientific name |
Fortiblatta cuspicolor Liang, Vršanský et Ren |
| status |
sp. nov. |
Fortiblatta cuspicolor Liang, Vršanský et Ren sp. nov.
( Figs.1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Holotype. CNU-B-NN-2006-006. Complete female with head, pronotum, abdomen, legs, terminalia and wings.
Additional material. Complete specimens: female: CNU-B-NN-2006-006, 0 0 7, 0 0 8, 0 0 9, 0 10, 0 70, 0 71, 073-1,073-2, 0 74, 0 76, 0 95, 108, 110, 213, 217, 258, 260, 492, 493, 494, 495, 496, 684-1, 684-2, 688-1, 688- 2, 701, 702; uncertain gender: 0 72, 0 75, 0 77, 0 78, 0 79, 0 80, 114, 125, 205, 215, 216, 218, 219, 235, 240, 250, 252, 261, 490, 491, 685, 703, 704, 705.
Type locality. Daohugou Village, Shantou Township, Ningcheng County, Inner Monglia, China.
Type horizon. Jiulongshan Formation, the Middle Jurassic.
Derivation of name. The name cuspicolor is after cuspis (Latin for cusp) and color - alluding to wing apex coloration.
Description. Large species (forewing length/width: 21.1–26.0mm/6.5–8.0mm; hind wing length/width: 21.0–24.0mm/7.0– 8.3mm).
Head prognathous, prolonged and narrow (length/width: 3–4.6mm / 2–2.5mm), unconcealed by pronotum, ( Figs.1 View FIGURE 1 a, 1b, 2a, 5b), the base of head with long setae ( Figs. 2 View FIGURE 2 a, 5b).
Pronotum vaulted, slightly elongate (length/width: 5.00– 5.88mm / 4.46 –5.59mm), with dark coloration at center and two slender color stripes along lateral margins ( Figs. 1 View FIGURE 1 a, 1b,).
Mandibles strong, with double sharp teeth. Labial palp 3-segmented, with 3rd segment being the longest, with a wide apex, and 2nd and 3rd segments with slender sparse chaeta (length of labial palpus: 2nd: 0.47–0.56mm; 3rd: 0.65–0.77mm). Maxillary palp 5- segmented, 3rd and 4th segments longer and wider than others, with stronger, denser setae (as compared to setae on labial palp). Segments of maxillary palps widened apically (length of maxillary palp segments: 2nd: 0.63mm; 3rd: 1.18–1.31mm; 4th: 1.11–1.32mm; 5th: 1.12–1.15mm) ( Figs. 2 View FIGURE 2 b, 5a).
Antenna long (longest preserved fragment long 16mm), filiform, multi-segmented, with more than 80 segments, the first segment length/width: 0.35mm / 0.28mm; antennal socket ovoid, slightly transverse (length/ width: 0.49mm / 0.58mm) ( Fig. 2 View FIGURE 2 b).
Intercalary veins well developed in all wings. Forewing long and narrow. Sc with 2–4 branches; R expanded, with 13–21 branches, basalmost branches simple; M divided basally into two straight branches, with total 5–12 veins at margin; CuA with 6–15 branches. Clavus long and narrow, anal veins with tertiary branches, with 7–15 veins at the margin. A diagonal kink in anal field ( Figs. 1 View FIGURE 1 a, 3b) is present. Hind wing with simple Sc; R1 with 4–9 branches and Rs with 10–15 branches; M branched basally in respect to Rs, and apically in respect to R1, Rs and CuA; CuA basalmost branches strong, CuP simple; A1 simple, A3 with secondarily branched veins. Reticulations present in CuA-CuP space, joined with intercalaries ( Figs. 1 View FIGURE 1 a, 1b).
Abdomen wide, body cuticle heavily sclerotised.
Fore leg with retained cursorial function, with very few spines, setae on tarsus longer and denser than femur and tibia. Coxa without setae and chaeta, twice wider as femur; trochanter forms a triangle; femur 1.4 times as long as coxa, with few spines; tibia little longer than femur, with sparse spines (when compared with femur); tarsus 5-segmented (length of tarsomeres: 1st: 1.03mm; 2nd: 0.40mm; 3rd: 0.36mm; 4th: 0.21mm; 5th: 0.30mm), the 4th segment smallest; terminal segment with a claw and arolium preserved. Length of leg segments: fore coxa, femur, tibia, tarsus, respectively 2.37–2.72mm; 3.43–3.70mm; 4.15–4.36mm; 2.81mm ( Figs. 2 View FIGURE 2 c, 5c). Mid leg with few spines on femur and tibia. Hind leg with denser spines on the tibia. Length of mid femora and tibiae: 5.35–5.75mm; 6.00mm. Length of hind femora and tibiae: 7.20–7.80mm; 10.25–11.28mm ( Figs. 2 View FIGURE 2 d, 5d).
Cerci very long (2.40–2.92mm, first segment length/width: 0.45mm / 0.19mm); composed of at least 10 segments, the last three segments longer and thinner than the others. Internalized are inner but also outer valves ( Figs. 2 View FIGURE 2 e, 5f–h).
Remarks: The setae on the maxillary palp are more sclerotized than the setae on the labial palp and leg (examined on specimen CNU-B-NN-2006-075) ( Figs. 2 View FIGURE 2 b, 5a), and the setae on the tarsus are longer, more densely distributed than the setae on the foreleg femur. This may be a result of different sensitivity or different manipulation function of P. mx vs. P. lb, and tarsus vs. femur ( Vršanský et al. 2001). It implies that this character might be essential for cockroaches in searching for and/or contacting with food and its environment.
Hind wing M obliquely with two branches, except for specimen CNU-B-NN-2006-261 RHW with five branches. The specimen has characteristics of the wings, body, and pronotum identical to those of the here described Fortiblatta cuspicolor , so we can place the specimen within the new species.
Fusion of veins is present in at least 13 of 61 wings (13/61) ( Fig. 4 View FIGURE 4 ), and on one wing (CNU-B-NN-2006- 492 RHW) ( Fig. 4 View FIGURE 4 g), three defromities are present. This ratio is the highest compared with the other localities, such as those in the Bohemian Massif (3 of 29), Carboniferous sediments of Germany (4/256), Bakhar in Monglia (5/500), Bon Tsagaan in Monglia (6/673), Montsec in Spain (2/121), Siberian Baissa (3/400) and Dobbertin in Germany (3/52), but comparable with the Tithonian of Shar-Teg in Mongolia (18/170) ( Vršanský 2005). Fused intercalary which were unknown before (CNU-B-NN-2006-008 LFW) ( Fig. 4 View FIGURE 4 h), and A1 joined with CuP (CNU-B-NN-2006-076 of RFW) ( Fig. 4 View FIGURE 4 e) probably represent heritable changes ( Vršanský 2005). In specimen CNU-B-NN-2006-261 RHW ( Fig. 4 View FIGURE 4 f), branch of M has apparent trend to connect with RS, but not join with it. It is an evidence for the action of compensatory (correlation of vein distance) mechanisms, which detect unusual development of the wing venation pattern.
Variability ( Tables 1–6 View TABLE 1 ). We analyzed wings including CNU-B-NN-2006-006LFW, 006RFW, 006LHW, 006RHW, 007LFW, 007RFW, 007LHW, 007RHW, 008LFW, 008LHW, 0 0 9 L FW, 009RFW, 010RFW, 070LFW, 070RFW, 071LFW, 071RFW, 072LFW, 073LFW, 073RFW, 074LFW, 076RFW, 077LFW, 078LFW, 079LFW, 095RFW, 108LFW, 108RFW, 108LHW, 114LHW, 213LFW, 213RFW, 213LHW, 216RFW, 217RFW, 217RHW, 219LFW, 235RFW, 235RHW, 260RFW, 261RFW, 261RHW, 490LFW, 490RFW, 491LFW, 492LFW, 494LFW, 495RHW, 496RFW, 684LFW, 684RFW, 684LHW, 684RHW.
A R1 M CuA R+M M+Cu R+Cu Sum Total
Minimum 8 13 5 6 22 16 2 29 42 Maximum 13 21 10 15 27 22 37 42 58 Median 10 16 9 10 24 20 28 35 49 Mode 9 14 10 9 24 20 28 34 46 Mean 10.16 16.11 8.53 9.74 24.63 19.32 26.84 35.32 49.11 Std. Error of Mean 0.31 0.47 0.39 0.47 0.28 0.43 0.86 0.63 0.81 Std. Deviation 1.38 2.05 1.71 2.05 1.21 1.86 3.76 2.75 3.53 CV. 13.63 12.74 20.08 21.06 4.92 9.60 14.01 7.79 7.18 A R1 M CuA R+M M+Cu R+Cu Sum Total
Minimum 7 13 4 9 21 14 24 31 44 Maximum 15 20 12 14 29 26 33 43 58 Median 12 17 8 11 24 20 28 36 52 Mode 12 19 8 12 24 22 30 33 49 Mean 11.58 16.53 8.37 10.84 24.89 20.21 28.37 36.74 51.84 Std. Error of Mean 0.453 0.57 0.41 0.33 0.63 0.64 0.65 0.78 0.88 Std. Deviation 1.95 2.50 1.80 1.46 2.72 2.78 2.83 3.40 3.83 CV. 16.86 15.14 21.52 13.49 10.95 13.76 9.98 9.25 7.40 The lowest CV value has the total vein number meeting the forewing margin (7.70 including Sc and A) (44–58). M with CV=20.54 (4–12) seems to be the most variable vein. Comparatively lower variation (CV= 8.42) in the fore margin (R+M) indicates improved flight abilities ( Vršanský 1997). Radial veins, important for flight as directing most power during the strike (at anterior margin of the wing), had less variable CV (13.90), which also indicates good flight. Limited variation in the anterior part of the wing suggests enhanced flight capabilities when compared with the representatives of the Spiloblattinidae Handlirsch, 1906 , Poroblattinidae Handlirsch, 1906 , Mylacrididae Scudder, 1886 , Caloblattinidae Vršanský et Ansorge, 2000 , Mesoblattinidae Handlirsch, 1906 , Blattulidae Vishniakova, 1982 ( Vršanský 2000) . The data support the hypothesis that the Raphidiomimidae were active flyers.
The variation in the hind wing (CV=7.33, excluding Sc and A), which bears the major weight during flying, is lower than that in the forewing (CV=8.65, excluding Sc and A), which is due to better vein distance correlation mechanisms, and also supports Raphidiomimidae were active in flight ( Vršanský 2003). Unlike Elisama tsaganica Vršanský, 1999 (hind wing CV=9.03), hind wing is more variable than the forewing caused by the possibility of the regulation of the vein strength in Blattulidae ( Vršanský 2003, Wang et al. 2007a). CV index on hind wing is also lower than Habroblattula drepanoides Wang, Liang et Ren, 2007 (CV=14.43) ( Blattulidae ) ( Vršanský 2003, Wang et al. 2007a).
The left and right wings differ in total number of veins (CNU-B-NN-2006-006LFW/ RFW: 50/52, 006LHW/ RHW: 54/48; 007LFW/ RFW: 48/49, 007LHW/ FHW: 50/52; 009LFW/ RFW: 49/54; 070LFW/ RFW: 51/44; 073LFW/ RFW: 58/57; 108LFW/ RFW: 50/54; 213LFW/ RFW: 51/57; 490LFW/ RFW: 49/46; 684LFW/ RFW: 48/50; 684LHW/ RHW: 54/ 52). All the particular CV index of left and right wings are shown in Tables 3, 4 View TABLE 4 , 5 View TABLE 5 , and 6. From these comparison data, it is clear that asymmetry between left and right wings was substantial, especially in the hind wings (including length, and number veins of RW and LW), which suggests the Raphidiomimidae were not such excellent fliers as some living cockroaches.
The present observation and the wing character analysis supports the hypothesis that the Raphidiomimidae originated from the representatives of the Caloblattinidae (which is presumed to be paraphyletic family with respect to Raphidiomimidae , Liberiblattinidae and some other extinct families) ( Vršanský 2002a). Both Raphidiomimidae and Caloblattinidae share strong synapomorphies such as wide abdominal segments, long palps, elongated wings with apparent intercalaries, diagonal fold in the forewing, hind wing with many reticulations in CuA-CuP space, R with R1 and RS abundantly branched, M weakly branched, CuA secondarily branched.
Grimaldi and Ross (2004) have assigned Raphidiomimula burmitica Grimaldi et Ross, 2004 , from the Burmese amber, to the Raphidiomimidae , but this taxon does not exhibit any of the apomorphic characters of the family ( Vršanský 2005, Gorochov 2006). The hypognathous prolonged head (wings are absent in that nymph) and short fore tibiae are indicative for the Caloblattinidae rather than Raphidiomimidae .
The proportion of femur and tibia of foreleg in Fortiblatta is different from the Late Jurassic Raphidiomima and Cameloblatta (in which the ratio is 1.06 (Vishniakova 1973)). Long and thin legs with foreleg tibia longer than femur in Fortiblatta imply possible six-legged locomotion when hunting. Tibiae displayed a tendency of shortening, to a level of almost the same as femur in the Late Jurassic, and became shorter than femur among mantises in the Upper Cretaceous (Vršanský, 2002). Eyes of new genus were located basally, and protected by a unique longitudinal ridge, which might have helped precise orientation during pursuit and protection of eyes. This structure has no equivalent in the Dictyoptera and is absent even in the Late Jurassic species found in Karabastau Formation.
Internalized outer valves of female ovipositor is an advanced character (Vršanský 2002), suggesting that eggs could not be laid deeply into the substrate or decaying wood. The inner valves of the new genus are shorter than in species from Karatau, suggesting advanced reproduction and eggs-laying associated with precursors of ootheca (Vršanský 2002, Anisyutkin and Gorokhov 2004)
Dark coloration of wings and body of Fortiblatta implies enhanced humidity and floral cover - in contrast to an open and barren environment ( Jarzembowski 1994).
The variability of the present species suggests Raphidiomimidae had active flight capability, better than some representatives of Blattulidae ( Blattula langfeldti (Geinitz, 1880) , Elisama parallela Vršanský, 2003 ), but according to the enhanced asymmetry, they were not excellent flyers.
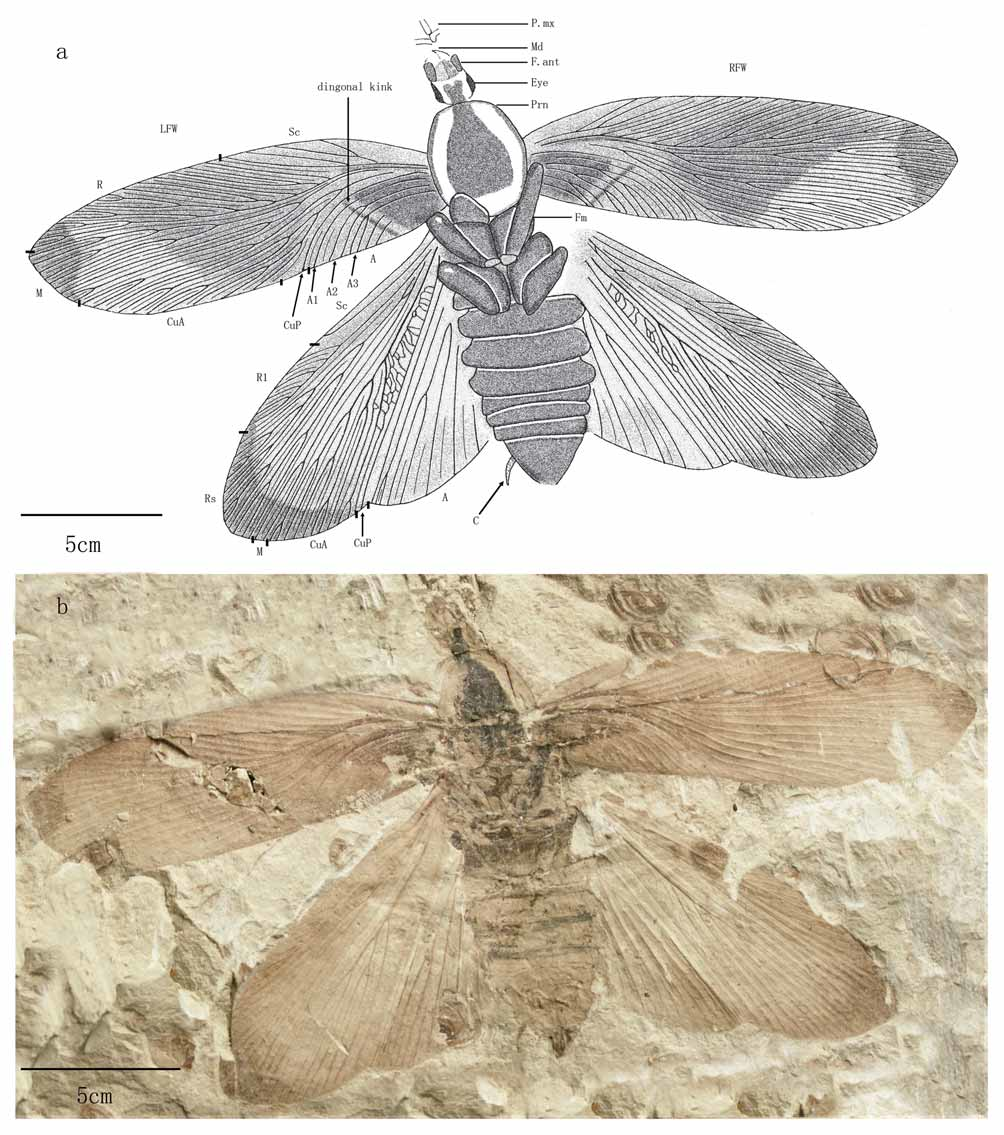
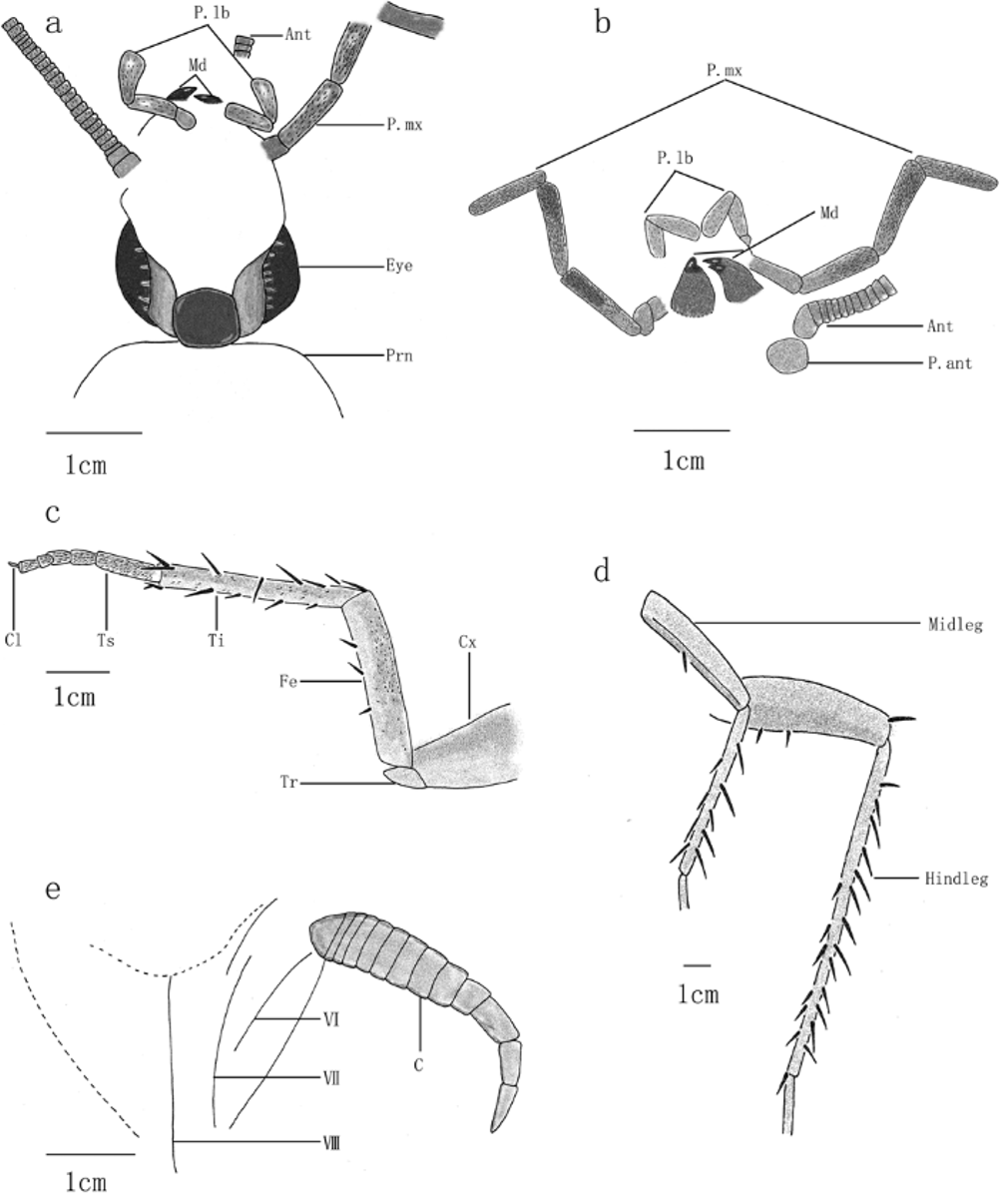




TABLE 4. Hind wing venation variability of Fortiblatta cuspicolor sp. nov.. Sum: Sc and A are excluded.
| R1 | Rs | R | M | CuA | R+M | M+Cu | R+Cu | Sum | |
|---|---|---|---|---|---|---|---|---|---|
| Minimum | 4 | 10 | 15 | 2 | 6 | 17 | 9 | 27 | 29 |
| Maximum | 9 | 15 | 24 | 5 | 12 | 26 | 18 | 34 | 36 |
| Median | 7 | 13 | 19 | 2 | 10 | 21 | 13 | 29 | 31 |
| Mode | 7 | 13 | 20 | 2 | 10 | 22 | 13 | 28 | 30 |
| Mean | 6.67 | 12.6 | 19.27 | 2.2 | 9.4 | 21.47 | 12.53 | 29.6 | 31.86 |
| Std. Error of Mean | 0.39 | 0.41 | 0.56 | 0.20 | 0.40 | 0.57 | 0.53 | 0.54 | 0.60 |
| Std. Deviation | 1.50 | 1.59 | 2.19 | 0.77 | 1.55 | 2.20 | 2.07 | 2.10 | 2.33 |
| CV. | 22.44 | 12.66 | 11.35 | 35.21 | 16.48 | 10.25 | 16.48 | 7.10 | 7.33 |
TABLE 5. Fortiblatta cuspicolor sp. nov. Left hind wing venation variability. Sum: Sc and A are excluded.
| R1 | Rs | R | M | CuA | R+M | M+Cu | R+Cu | Sum | |
|---|---|---|---|---|---|---|---|---|---|
| Minimum | 4 | 10 | 15 | 2 | 6 | 17 | 9 | 27 | 29 |
| Maximum | 8 | 15 | 20 | 2 | 12 | 22 | 15 | 31 | 33 |
| Median | 6.5 | 13 | 19 | 2 | 10 | 21 | 13 | 28.5 | 30.5 |
| Mode | 7 | 13 | 20 | 2 | 10 | 22 | 13 | 31 | 33 |
| Mean | 6.13 | 12.5 | 18.63 | 2 | 9.25 | 20.63 | 12.25 | 28.88 | 30.88 |
| Std. Error of Mean | 0.48 | 0.53 | 0.59 | 0 | 0.65 | 0.60 | 0.65 | 0.58 | 0.58 |
| Std. Deviation | 1.36 | 1.51 | 1.69 | 0 | 1.83 | 1.69 | 1.83 | 1.64 | 1.64 |
| CV. | 22.14 | 12.09 | 9.00 | 0 | 19.81 | 8.17 | 14.96 | 5.69 | 5.32 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.