
Geonoma macrostachys Martius (1823: 19)
Henderson, Andrew, 2011, A revision of Geonoma (Arecaceae), Phytotaxa 17, pp. 1-271 : 93-95
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.17.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5609049 |
|
persistent identifier |
https://treatment.plazi.org/id/FA4887FA-2442-BD19-FF37-DC3B1A70A4CA |
|
treatment provided by |
Plazi |
|
scientific name |
Geonoma macrostachys Martius (1823: 19) |
| status |
|
36. Geonoma macrostachys Martius (1823: 19) View in CoL . Taenianthera macrostachys (Martius) Burret (1930a: 268) .
Type: BRAZIL. Amazonas: Rio Japurá, no date, C. Martius s.n. (holotype M!).
Geonoma acaulis Martius (1823: 18) View in CoL . Taenianthera acaulis (Martius) Burret (1930a: 269) . Geonoma macrostachys var. acaulis (Martius) Henderson (1995: 274) View in CoL . Type: COLOMBIA. Amazonas: “Rio Negro ad montem Cupati” [Río Caquetá, Cerro Yupatí], no date, C. Martius s.n. (holotype M!).
Geonoma tamandua Trail (1876: 323) View in CoL . Taenianthera tamandua (Trail) Burret (1930a: 268) . Type: BRAZIL. Amazonas: Rio Javari, São Antonio de Boa Vista 4 December 1874, J. Trail 976/CLXXXIII (holotype K!, isotypes GH!, P!).
Geonoma acaulis subsp. tapajotensis Trail (1876: 324) View in CoL . Geonoma tapajotensis (Trail) Drude (1882: 508) View in CoL . Taenianthera tapajotensis (Trail) Burret (1930a: 269) . Type: BRAZIL. Pará: Aramanahy, Rio Tapajós, 10 January 1874, J. Trail 1017/IX (holotype K!).
Taenianthera oligosticha Burret (1931a: 201) View in CoL . Type: PERU. Loreto: Río Nanay, May-June 1929, L. Williams 737 (holotype F!).
Geonoma atrovirens Borchsenius & Balslev in Borchsenius et al. (2001: 342) View in CoL . Type: ECUADOR. Napo: Jatun Satcha Biological Field Station, SE of Mishualli, ca. 400 m, 2 October 1995, H. Balslev et al. 6430 (holotype AAU!, isotype QCA, n.v.), synon. nov.
Geonoma supracostata Svenning in Borchsenius et al. (2001: 344) View in CoL . Type: ECUADOR. Napo: E of Yasuní Scientific Station, Yasuní National Park, 0°40’S, 76°23’W, 23 March 1995, J.-C. Svenning 148 (holotype AAU!), synon. nov.
Geonoma ecuadoriensis Henderson, Borchsenius & Balslev (2008: 192) View in CoL . Type: ECUADOR. Napo: carretera Hollin- Loreto-Coca, km 40, entre Río Guamaní y Río Pucuno, 0°40’S 77°38’W, 1200 m, 11 December 1987, D. Neill, W. Palacios & C. Cerón 8073 (holotype NY!, isotypes AAU!, MO n.v.), synon. nov.
Plants 1.3(0.3–3.5) m tall; stems 0.3(0.1–1.0) m tall, 1.9(1.1–2.8) cm in diameter, solitary or clustered, not cane-like; internodes 0.2(0.1–0.4) cm long, not scaly. Leaves 9(3–15) per stem, undivided or irregularly pinnate, not plicate, bases of blades running diagonally into the rachis; sheaths 13.2(2.3–28.0) cm long; petioles 35.5(0.0–137.5) cm long, drying green or yellowish; rachis 46.1(9.3–193.0) cm long, 3.7(1.3–9.8) mm in diameter; veins raised and rectangular in cross-section adaxially or not raised or slightly raised and triangular in cross-section adaxially; pinnae 3(1–16) per side of rachis; basal pinna 24.5(11.0–70.0) cm long, 4.4(0.3–32.0) cm wide, forming an angle of 39(3–100)° with the rachis; apical pinna 20.8(7.2–44.5) cm long, 8.8(1.4–28.0) cm wide, forming an angle of 31(8–77)° with the rachis. Inflorescences unbranched; prophylls and peduncular bracts ribbed with elongate, unbranched fibers, both bracts tubular, narrow, elongate, closely sheathing the peduncle, more or less persistent; prophylls 13.0(5.0–33.0) cm long, not short and asymmetrically apiculate, the surfaces not ridged, without unequally wide ridges; peduncular bracts 23.2(10.7–34.5) cm long, well-developed, inserted 1.1(0.1–22.6) cm above the prophyll; peduncles 59.2(19.2–128.5) cm long, 2.9(0.7–6.8) mm in diameter; rachillae 1, 14.1(4.2–31.0) cm long, 5.87(1.7–12.3) mm in diameter, the surfaces without spiky, fibrous projections or ridges, drying brown or yellow-brown, without short, transverse ridges, not filiform and not narrowed between the flower pits; flower pits spirally arranged, glabrous internally; proximal lips with a central notch before anthesis, often the two sides of the notch overlapping, not recurved after anthesis, not hood-shaped; proximal and distal lips drying the same color as the rachillae, not joined to form a raised cupule, the proximal lip margins overlapping the distal lip margins; distal lips well-developed; staminate and pistillate petals emergent, valvate throughout or not emergent, not valvate throughout; staminate flowers deciduous after anthesis; stamens 6; thecae diverging or not diverging at anthesis, inserted onto poorly to well-developed, non-split, jointed connectives, connectives when well-developed alternately long and short; anthers short at anthesis, remaining straight and parallel; non-fertilized pistillate flowers deciduous after anthesis; staminodial tubes lobed at the apex, the lobes spreading at anthesis, acuminate, those of non-fertilized pistillate flowers not projecting and persistent after anthesis; fruits 8.9(6.2–13.7) mm long, 6.5(4.7–9.8) mm in diameter, the bases without a prominent stipe, the apices not conical, the surfaces not splitting at maturity, without fibers emerging, bumpy from the numerous, subepidermal, tangential, short fibers present, these coming to a point at fruit apices; locular epidermis with operculum, smooth or sculpted and then usually also with a raised, meridional ridge, without pores.
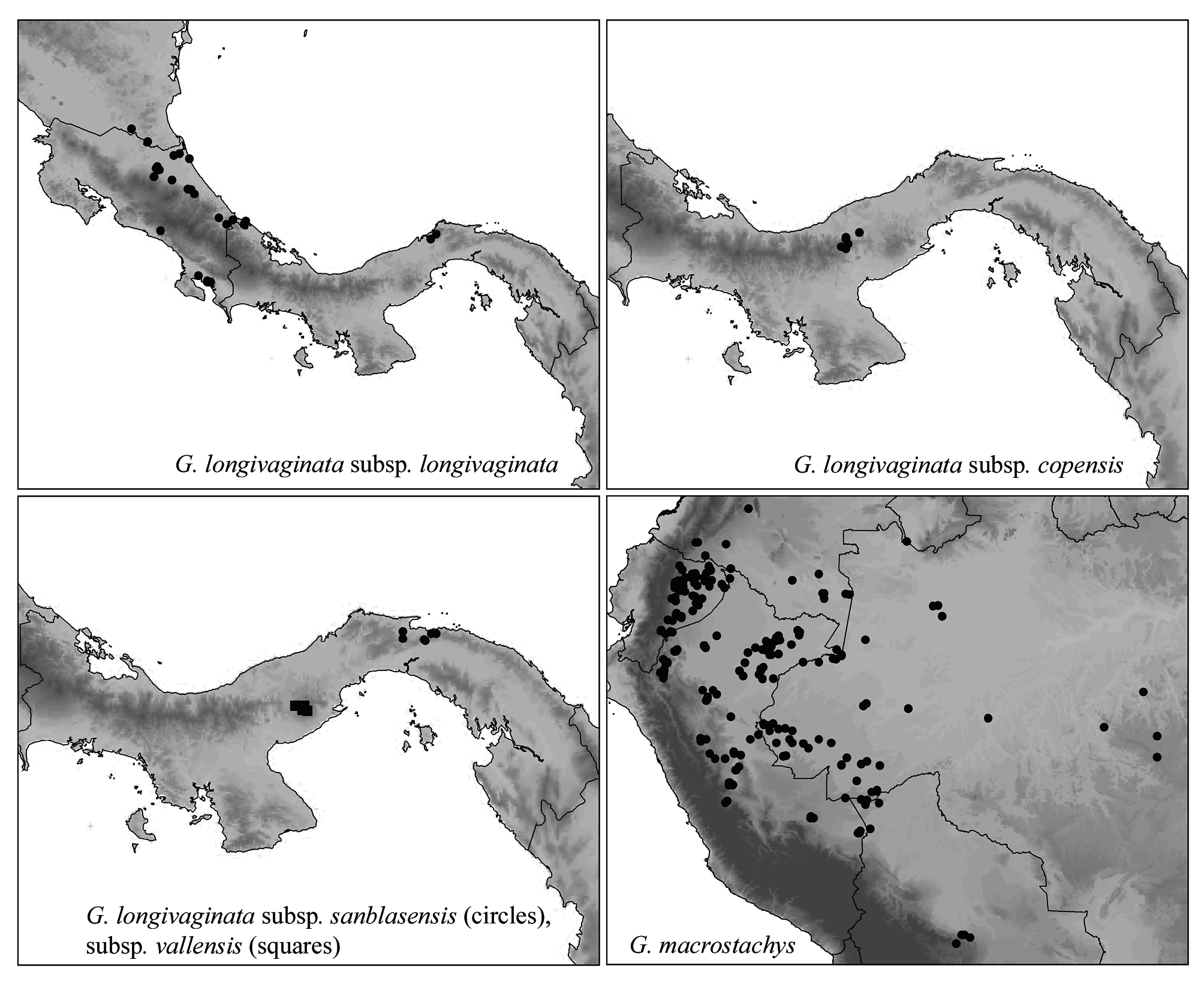
Distribution and habitat:— From 2°45’N– 17°50’S and 55°00’– 78°30’W in the western Amazon region in Venezuela, Colombia, Ecuador, Peru, and Brazil at 443(75–1800) m elevation in lowland or montane rainforest ( Fig. 24 View FIGURE 24 ).
Taxonomic notes:— Geonoma macrostachys is the most variable species in the whole genus, and most difficult taxonomically. It has been dealt with differently by both Wessels Boer (1968) and Henderson et al. (1995). However, no satisfactory treatment is possible based on herbarium specimens because variation is so complex and there is little geographic disjunction. This is discussed in some detail in the section on Infraspecific Variation. In this treatment a broad approach is taken and only one species recognized with various morphotypes, as discussed below.
Subspecific variation:— Five traits vary within this species (stem branching, leaf division, adaxial veins, petals, locular epidermis sculpting). Excluding two of these (stem branching, leaf division), and one for which there are few data (locular epidermis sculpting), the state combinations of the other two traits (adaxial veins, petals) divide the species into three subgroups (raised veins/non-exerted petals; non-raised veins/non-exerted petals; non-raised veins/exerted petals). There is no geographic discontinuity between any of these trait subgroups. Geonoma macrostachys is distributed widely in the central and western Amazon region. These three subgroups are not consistent. Within each there is much quantitative variation such that there are no clearly defined subgroups. Furthermore, there is unexplained variation in staminate flowers.
All specimens with staminate flowers are scored as having the thecae inserted onto poorly to welldeveloped, non-split, jointed connectives, when well-developed the connectives alternately long and short. However, there appear to be two different kinds of connective, poorly-developed and well-developed, and these two can occur together at the same locality. For example, in Ecuador (Napo), on eastern Andean slopes at 1200 m near the Río Guamaní, Neill 8073 has poorly-developed connectives and occurs near to, and is apparently identical with Neill 8622, which has well-developed connectives. In Peru (Madre de Dios) at low elevations in Amazon forests near Puerto Maldonado, Gentry 68666 has poorly-developed connectives, and is apparently identical with Timana 1588 which has well-developed connectives. Lacking staminate flowers, it is not possible to distinguish two kinds of connective. This problem cannot be solved with the material available. Although G. macrostachys is extremely variable, a number of morphotypes can be distinguished.
In the western Amazon region in Colombia, Ecuador, Peru, and Brazil, with an outlying population in southern Peru, there is a morphotype ( macrostachys ) with usually undivided leaves, non-raised veins, and 1(1–4) pinnae with narrow basal angles of 10(3–40)°. Staminate flowers have well-developed connectives, jointed at the apices of the filament, and alternately long and short. Specimens of this morphotype lacking staminate flowers and having undivided leaves are difficult to distinguish from other morphotypes (e.g., a trovirens). One specimen (Díaz 7058) from Peru has an exceptionally long interbract distance of 22.6 cm, and the prophyll is longer than the peduncular bract. The mean interbract distance of macrostachys without this specimen is 0.6 cm and the prophyll is always shorter than the peduncular bract. Specimens from Amazonas in Peru are particularly variable. Some specimens have wider leaves and almost sigmoid venation and others have unusually large leaves. The type of G. macrostachys is of this morphotype.
In the western and central Amazon region in Venezuela, Colombia, Ecuador, Peru, and Brazil there is a morphotype ( acaulis ) having pinnate leaves, non-raised adaxial veins, 5(3–9) pinnae with wide basal angles of 61(40–90)°, and staminate flowers with poorly developed connectives. All the specimens assigned to this morphotype have staminate flowers, but there are similar specimens, lacking staminate flowers, that are assigned to the tapajotensis morphotype (see below). One acaulis specimen (Balick 1140) from northwest of Iquitos has pinnae with raised veins. This occurs in an area without other acaulis but in which G. brongniartii is abundant.
The types of G. acaulis and G. ecuadoriensis are of the acaulis morphotype. The original description and illustration of G. acaulis by Martius (1823) showed staminate flowers with poorly-developed connectives. However, the illustration in Drude (1882), presumably using a different specimen (possibly a Trail specimen) than that of Martius (1823), shows the staminate flowers with well-developed, non-split connectives, these jointed at the apices of the filament. Because of this, Geonoma acaulis was subsequently interpreted as having ‘ Taenianthera - type ’ staminate flowers with well-developed connectives ( Wessels Boer, 1968; Henderson, 1995; Borchsenius et al., 2001).
In the western and central Amazon region in Colombia, Ecuador, Peru, Bolivia, and Brazil, with an outlying population in Bolivia, there is a morphotype ( tapajotensis ) that is similar to the acaulis morphotype in its pinnate leaves with non-raised veins and 4(2–10) pinnae with wide basal angles of 58(27–100)°, but has well-developed connectives. The types of G. acaulis subsp. tapajotensis and Taenianthera oligosticha are of this morphotype.
Some specimens (large-size morphotype), similar to the tapajotensis morphotype, from central Peru and Brazil (Acre), are larger in size but overlap with normal–sized specimens. They have pinnate leaves with nonraised veins, 4(3–9) pinnae with wide basal angles of 51(20–77)°, and well-developed connectives. They are extremely variable and some, apart from their non-raised veins, resemble the large-raised morphotype (see below). These large-size specimens occur sympatrically with large size specimens of G. brongniartii .
In the western Amazon region of Ecuador and adjacent Peru and Colombia there is a morphotype ( atrovirens ) with undivided leaves with non-raised veins with narrow basal angles of 7(5–10)°. On specimen labels the leaves are reported to be dark green or ‘black–green’. Connectives are almost intermediate between well-developed and poorly-developed ones. However, two, outlying specimens from Colombia (Galeano 1887, 2096) have poorly-developed connectives. Without staminate flowers it is difficult to distinguish this morphotype from the morphotypes m acrostachys or tamandua . The type of G. atrovirens is of this morphotype.
In the western Amazon region of Colombia, Ecuador, Peru, and Brazil there is a morphotype ( tamandua ) with exceptionally long rachis and undivided leaves with non-raised veins with narrow basal angles of 7(3– 17)°. The leaves are described on labels as dark green, blue green, ‘almost black’, ‘nearly black’, or ‘dark black-green’. In leaf color this morphotype resembles the atrovirens morphotype, but has well-developed connectives. Specimens come from two areas—Ecuador and adjacent Colombia, and Peru and Brazil. There is no difference between these two populations except that specimens from Ecuador and adjacent Colombia have wider rachillae. The type of G. tamandua is of this morphotype.
In the western Amazon region of Ecuador and Peru there is a morphotype ( supracostata ) having undivided or pinnate leaves with raised adaxial veins and 2(1–4) pinnae with wide basal angles of 26(10–48)°. Staminate flowers have well-developed connectives. The type of G. supracostata is of this morphotype.
In the western Amazon region of Ecuador and Peru there is a morphotype (large-raised) with usually divided leaves with raised adaxial veins and 7(1–16) pinnae and narrow basal pinna angles of 10(9–12)°. Staminate flowers have well-developed connectives. Specimens of this morphotype lacking staminate flowers and/or fruits are difficult to distinguish from specimens of G. multisecta and G. schizocarpa .
In the Peruvian Amazon and adjacent parts of Colombia there is a morphotype (grandiflora) with nonraised adaxial veins, 4(3–7) pinnae per side of the rachis, and wide basal angles of 66(44–90)°. The staminate and pistillate petals are emergent and valvate throughout at anthesis, and they tend to be larger than those of other specimens. The proximal lip of the flower pit is also more pronounced. One specimen (Gentry 54536) has 11 stamens.
| C |
University of Copenhagen |
| M |
Botanische Staatssammlung München |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Geonoma macrostachys Martius (1823: 19)
| Henderson, Andrew 2011 |
Geonoma ecuadoriensis
| Henderson, A. & Borchsenius, F. & Balslev, H. 2008: 192 |
Geonoma atrovirens Borchsenius & Balslev in Borchsenius et al. (2001: 342)
| Borchsenius, F. & Balslev, H. & Svenning, J. - C. 2001: 342 |
Geonoma supracostata Svenning in Borchsenius et al. (2001: 344)
| Borchsenius, F. & Balslev, H. & Svenning, J. - C. 2001: 344 |
Taenianthera oligosticha
| Burret, M. 1931: 201 |
Geonoma tamandua
| Burret, M. 1930: 268 |
| Trail, W. 1876: 323 |
Geonoma acaulis subsp. tapajotensis
| Burret, M. 1930: 269 |
| Drude, O. 1882: 508 |
| Trail, W. 1876: 324 |
Geonoma acaulis
| Henderson, A. 1995: 274 |
| Burret, M. 1930: 269 |
| Martius, C. 1823: 18 |