
Gobiosoma alfiei, Van Tassell, James L., Joyeux, Jean-Christophe, Macieira, Raphael Mariano & Tornabene, Luke, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4007.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:F6A8C8AF-5A2B-4F07-9679-5F0E026E776F |
|
DOI |
https://doi.org/10.5281/zenodo.6108038 |
|
persistent identifier |
https://treatment.plazi.org/id/86A723DC-5CE9-4651-9134-B2BFA90A6E70 |
|
taxon LSID |
lsid:zoobank.org:act:86A723DC-5CE9-4651-9134-B2BFA90A6E70 |
|
treatment provided by |
Plazi |
|
scientific name |
Gobiosoma alfiei |
| status |
sp. nov. |
Gobiosoma alfiei View in CoL sp. nov. Joyeux & Macieira
Alfie’s goby
Common name: Alfie’s goby Nome comum (Portuguese): Amboré do Alfie Figs. 6 View FIGURE 6 , 7 View FIGURE 7 , 8 View FIGURE 8
Gobiosoma hemigymnum (Eingenmann & Eingenmann 1888) View in CoL (misidentified; error in authors’ names). Joyeux et al., 2009: 64 – 66 (cephalic head pores pattern, habitat); Macieira & Joyeux, 2011: 311 (listed).
Gobiosoma hemigymnum ( Eigenmann & Eigenmann 1888) View in CoL (misidentified). Macieira & Joyeux, 2011: 311 (listed).
Gobiosoma spilotum ( Ginsburg 1939) View in CoL . (misidentified) Rocha et al., 1998: 564 (listed).
Gobiosoma parri Ginsburg 1933 View in CoL . (misidentified) Gomes et al., 2014: 1001 –1003 (larvae in a Bahian estuary).
Gobiosoma View in CoL sp. Joyeux et al., 2004: 1281, 1283 (larvae in an Espírito Santo´s estuary)
Material examined: Types. Holotype: CIUFES 3114, 20.6 mm TL, male, J.-C. Joyeux, R.M. Macieira & C.R. Pimentel, 28 October 2007, tidepool at Praia dos Castelhanos, Anchieta, Espírito Santo, Brazil, 20°50'S 40°37'W.
Paratypes: PARAÍBA, BRAZIL: UFPB 4296, 1, P.B. Feitoza & L.A. Rocha, 0 8 April 1999, Taci Grande de Cadebelo, Praia Formosa, Cabedelo; UFPB 4882, 1, R.S. Rosa, 25 September 1992, Ponta Seixas, João Pessoa.
BAHIA, BRAZIL: CIUFES 0 544, 2, A. Carvalho-Filho, 0 8 September 2007, tidepool at Praia do Forte, Mata de São João.
ESPÍRITO SANTO, BRAZIL: Collected from tidepools at Praia dos Castelhanos, Anchieta, 20°50'S, 40°37'W: AMNH 258026, 5, CIUFES 0 103, 1, (c&s), CIUFES 0 106, 1, c&s, R.M. Macieira & E.R.S. Almeida, 21 August 2005; CIUFES 0 130, 1, c&s, CIUFES 0 131, 2, c&s, R.M. Macieira, J.L. Gasparini & J.-C. Joyeux, 15 November 2005; UF 187658, 3, R.M. Macieira & J.-C. Joyeux, 24 August 2006; UF 187659, 3, R.M. Macieira, J.- C. Joyeux, J.L. Gasparini, C.R. Pimentel & P. Sant'Ana, 18 March 2007; AMNH 258025, 2, R.M. Macieira, J.-C. Joyeux, J.L. Gasparini, C.R. Pimentel & P. Sant'Ana, 18 March 2007; AMNH 258029, 1, CIUFES 0 263, 1, R.M. Macieira, J.L.Gasparini, C.R. Pimentel & P. Sant'Ana, 17 June 2007; AMNH 258027, 1, R.M. Macieira & J.-C. Joyeux, 14 May 2006; CIUFES 0 274, 4, R.M Macieira, J.-C. Joyeux, J.L. Gasparini, P. Sant'Ana & C.R. Pimentel, 15 February 2006; MZUSP 112485, 17, CIUFES 0 525, 5, collected with holotype, J.-C. Joyeux, R.M. Macieira & C.R. Pimentel, 28 October 2007; ZUEC 9030, 6, A. Carvalho-Filho, R.M. Macieira & C.R. Pimentel, 30 August 2008.
ESPÍRITO SANTO, BRAZIL: Additional locations. CIUFES 0 201, 4, J.-C. Joyeux & R.C. Smarzaro, 29 November 2005, tidepool at Praia Escondida, Aracruz; CIUFES 0 646, 4, R.G. Santos, 28 November 2007, collected during algae sampling, Praia de Santa Helena, Vitória; CIUFES 1449, 2, J.L. Gasparini & S. Vogel, 0 3 November 1994, Santa Cruz, Aracruz; CIUFES 2006, 1, CIUFES 2007, 1, CIUFES 2008, 1, CIUFES 2009, 1, R.M. Macieira; J.L. Gasparini & J-C. Joyeux, 23 November 2010, tidepool at Barra do Riacho, Aracruz; CIUFES 2245, 1, R.M. Macieira, H.T. Pinheiro & L.B.C. Xavier, 27 September 2011, Manguinhos, Serra.
Non-type material: CEARÁ, BRAZIL: GEA.ICT 0 0 307, 1, R.R.S. Oliveira & F.S. Machado, 0 8 May 2012, tidepool, Iparana, 03°41´08.4S, 38°38´04.9W.
RIO GRANDE DO NORTE, BRAZIL: GEA.ICT 0 0 301, 2, R.R.S. Oliveira & F.S. Machado, 0 4 May 2012, tidepool, São Miguel do Gostoso, 05°06´05.9S, 35 ° 41´57.5W.
PARAÍBA, BRAZIL: UFPB 4023, 7, P.B. Feitoza et al., 0 4 November 1998, reefs at Praia do Poço, south of Areia Vermelha, Cabedelo; UFPB 4258, 1, P.B. Feitoza et al., 25 July 1998, tidepools north of Areia Vermelha, Cabedelo; UFPB 4610, 3, M.A. Guimarães et al., 18 July 1982, Ponta do Guajura.
PERNAMBUCO, BRAZIL: UF 141848, 1, T. Roberts, 31 March 1968, tidepool in front of LACIMAR (Laboratório de Ciências do Mar), Piedade; UF 141867, 6, T. Roberts, 0 3 May 1968, large open sandy tidepool beside old sewage canal at Paria (?), Recife.
ALAGOAS, BRAZIL: UF 19907, 3, N. Menezes, 12 December 1973, Ponta Verde, Maceió, 9.66408 S - 35.69463W.
BAHIA, BRAZIL: USNM 274857, 3, N.A. Menezes et al., 21 March 1985, Coroa Vermelha reef, Prado, 17°06' S, 39°11' W, depth 0–2.5 feet; CIUFES 1385, 1, A. Carvalho-Filho, no date, Praia do Forte, Mata de São João; GEA.ICT 0 1728, 1, R.R.S. Oliveira & F.S. Machado, 19 April 2012, tidepool, Cabralia, 16°19´53.6 S, 39°00´02.3W.
ESPÍRITO SANTO, BRAZIL: Collected from tidepools at Praia dos Castelhanos, Anchieta, 20°50'S, 40°37'W; CIUFES 0 0 39, 1, J.L. Van Tassell, J.-C. Joyeux & R.M. Macieira, 18 February 2004; CIUFES 0 107, 1, R.M. Macieira & E.R.S. Almeida, 21 August 2005; CIUFES 0 132, 1, CIUFES 0 271, 1, R.M. Macieira, J.L. Gasparini & J.-C. Joyeux, 15 November 2005; CIUFES 0 217, 1, R.M. Macieira, J.-C. Joyeux, J.L. Gasparini & P. SantAna, 0 4 November 2006; CIUFES 0 265, 5, R.M. Macieira, J.L. Gasparini, C.R. Pimentel & P. SantAna, 17 June 2007; CIUFES 0 272, 2, R.M. Macieira, J.-C. Joyeux, J.L. Gasparini, C.R. Pimentel & P. SantAna, 15 February 2006; AMNH 258024, 22, J.-C. Joyeux & R.M. Macieira, 28 October 2007; UF 184467, 23, J.-C. Joyeux & R.M. Macieira, 28 December 2007.
ESPÍRITO SANTO, BRAZIL: Additional locations. CIUFES 1118, 1, tidepool, Praia Escondida, Aracruz, J.- C. Joyeux & R.C. Smarzaro, 25 March 2005; CIUFES 2245, 1, tidepool, Praia de Manguinhos, Serra, R.M. Macieira, H.T. Pinheiro & L.B.C. Xavier, 27 September 2011; SIO 14–42, 8, R.M. Macieira & J.L. Gasparini, 0 2 June 2011, tidepool, Barra do Riacho, Aracruz; ZUEC 3179, 1, J.L. Gasparini & E.L.C. Dall'Orto Filho, 22 April 1987, between Ilha Galheta de Dentro and Ilha do Boi, Vitória; ZUEC 3178, 1, J.L. Gasparini & M.B. Collodetti, 0 6 February 1988, Ilha Galheta de Dentro, Vitória; CIUFES 130031, 1, CIUFES 130036, 14, E.C. Perrone, E.B. Santana, P.S.B. Barbosa & J.L. Helmer, 12 September 1983, tidepool, Praia de Capuba, Serra; CIUFES 130346, 1, C. Zamprogno, 15 September 1986, tidepool, Praia da Baleia, Serra;
Unsettled larvae from ichthyoplankton samples: IPK 0 268, 7, IPK 0 269, 6, IPK 0 270, 3, J.-C. Joyeux & B.B. Pereira, 24 September 1999, Canal da Passagem, Vitória.
Diagnosis. A species of Gobiosoma distinguished from other members of the genus by the following characters: body scaled from caudal peduncle to base of pectoral fin, scales in a wedge shape to under posterior of first dorsal-fin base, then continuing anteriorly in a row of 3–4 scales, row slightly tapering anteriorly; a naked area extending from mid-pectoral-fin base to 4th—11th ray of second dorsal fin (rarely extending further) and from the mid-pectoral-fin base to 3rd—9th anal-fin ray (rarely extending further); upper jaw 21.5% in head length; lateral scales modally 29; individuals 15 mm SL or larger with 24+ scales; pectoral fin 18 (17–20); maximum size to 27.8 mm SL.
General morphology. Morphometrics and meristics of the holotype and paratypes are given in Table 4 View TABLE 4 . Body cylindrical, elongate; head rounded in profile; eyes dorsolateral 6.7–9.6% in SL; maxilla generally extending posteriorly to a vertical ventral to mid-pupil; upper jaw length 4.0–9.6% in SL; anterior naris an elongate tube without a flap; posterior naris a raised rim to short tube; very small, wart-like barbel in front of eye, near anterior naris; males with a triangular, wide, flattened urogenital papilla; female papilla, slightly tapering, not flattened, with large opening at end; juveniles with urogenital papilla similar to that of males but shorter, less flattened and rounder at terminus.
Fins. Counts for holotype followed by (range; mode). First dorsal VII (VI–VII, single specimen with VI; VII), no elongate or filamentous spines; first dorsal connected to second dorsal by low membrane; second dorsal fin I,10 (I, 8–I,11; I,11), not reaching caudal fin when adpressed; anal fin I,9 (I, 5–I,9; I,9), not reaching caudal fin when adpressed; pectoral counts for left fin followed by right, 18/18 (17–20/17–20; 18/18), fin ovate, extending to anus when adpressed; caudal fin round, segmented rays 15 (14–17; 17), branched rays 14 (11–15; 14); pelvic fin I,5 (I,5; I,5), oblong to ovate, extending 3/4 distance to anus when adpressed.
Scales ( Fig. 4 View FIGURE 4 B). Counts for holotype followed by (range; mode). Lateral scales rows 29 (3–33; 29); head, nape, abdomen, prepelvic region and belly without scales; scales ctenoid, with pigmented posterior edge; body scaled generally from caudal peduncle to base of pectoral fin, scales in a wedge shape to under posterior of first dorsal-fin base, then continuing forward in a row, reducing to 3–4 scales under pectoral-fin base; a naked area extending diagonally from mid-pectoral-fin base to (or posterior to) the 4th ray of second dorsal fin (generally ending before posterior of D2 base) and from the mid-pectoral-fin base to (or past) the 3rd anal-fin ray (generally ending before end of anal fin base); four basicaudal scales extending on to caudal fin base, the dorsal and ventralmost with elongate ctenii ( Fig. 4 View FIGURE 4 E). Juveniles show scale formation that progresses from posterior to anterior, adding scales at the anterior, dorsal and ventral edges of the wedge; basicaudal scales are present in settlement larvae ( Fig. 9 View FIGURE 9 ).
Teeth. Based on five cleared and stained specimens. Females: Upper jaw with 3–4 rows anteriorly, reduced to a single row posteriorly, extending nearly to end of premaxilla; all teeth conical pointed and recurved. Outer row of teeth large recurved, conical and evenly spaced; no canine teeth present; lower jaw with 3–4 rows anteriorly, reduced to 1 row posteriorly, outer and inner row slightly larger than central rows, teeth conical with recurved tip; no canine teeth present; teeth extending to terminus of dentary. Males: Upper jaw with 3–4 rows anteriorly reduced to a single row at posterior, extending nearly to terminus of premaxilla; all teeth conical, pointed and recurved; outer row with large recurved conical teeth, evenly spaced; no canine teeth present; lower jaw with 3–4 rows at anterior, outer row slightly larger, middle rows small, closely packed, inner row with 3–4 large recurved canine teeth along anterior third of jaw; teeth extending to terminus of dentary.
Lateral-line system ( Fig. 8 View FIGURE 8 ). Cephalic lateral line system with pores B', C(s), D(s), E, F, H' and posterior oculoscapular pores K', L'; preopercle canal with pores M', N and O'; supraorbital canals fused between the eyes with only single canal present; small individuals (<17 mm TL) occasionally missing one or more of the following pores: K, L or M, N.
Sensory papillae in a transverse pattern ( Fig. 8 View FIGURE 8 ); row 1 extending from orbit to row d; rows 2 and 3, not reaching orbit; row 5 s from orbit to near anterior of b, but not connecting to b; row 6 not reaching row b; row 7 represented by 1 papillae (rarely 2); row b short, ending anteriorly before row 5 s; row d continuous; row 5 i under b and extending below level of d by 2–3 papillae. Dorsal row g with 3–4 papillae; row o reduced to 1 papilla; row m not observed and row n extending from level of pore F, medially to just past pore E.
Color in life ( Fig. 6 View FIGURE 6 ). Background color of head and body brownish to olive-green, occasionally with a golden tint. Body with eight bars along body from under anterior of first dorsal fin to caudal fin; each bar of mottled brown with a light (whitish) thin central area; edges of bars not well defined, with indistinct wavy edges; bars not extending onto the belly and often the bars are not evident in males. First bar located below the anterior of first dorsal fin (D1), second under posterior of D1, third between D1 and D2, 4th through 6th equally spaced under D2, 7th on caudal peduncle and the 8th, a thin bar, at the posterior of the caudal peduncle. A series of single dark spots along lateral midline (spots occasionally double) located in the middle of each bar; last dot, of 8th bar, often as a forwardpointing triangle. Head with three spots located in a line between the posterior of the eye and the posterior edge of the opercle; first located behind eye, second above posterior edge of preopercle and third above posterior edge of opercle. Side of head with two bars; first from the anteriorventral edge of the eye to the upper lip, extending over the lip and occasionally onto the lower lip, bar not always distinct in appearance; a second more distinct bar from mid-ventral orbit, vertically to and past posterior of jaw, bar with a short posteriorly directed extension, below midpoint of bar. Posterior edge of preopercle dark, along most of its length. Opercle with dark anterior edge, not extending the full length. Dorsal fins with horizontal bands, continuous in males, pigment in females restricted to elements. First dorsal fin with three oblique dark bands, the lower two bands coalescing, at their posterior, with the first and second dark bars of the body; upper band located along dorsal edge of fin. Second dorsal fin with 4 oblique bands, first three coalescing, at their posterior ends, with bars of body; fourth band near dorsal margin of fin; bands poorly defined. Anal fin with two poorly defined horizontal bands along its dorsal and ventral edges. Pectoral fin with two diffuse dark spots (sometimes crescent shaped), on dorsal and ventral base of fin rays; remainder of the fin transparent. Pelvic fin dusky with melanophores concentrated between elements. Caudal fin transparent with a few scattered melanophores, forming light vertical bars in both sexes. The species is dimorphic in pigment pattern. In breeding males the head and the anterior part of the trunk is yellowish orange, body color darker and banding less obvious; in females the head coloration is similar to that of the body.
Preserved color ( Fig. 7 View FIGURE 7 ). Preserved specimens retain their distinct color pattern and dimorphism persists (7 years in alcohol 70%) despite fading; the brightest colors (reddish brown and golden yellow) vanish leaving only the underlying melanophores or unpigmented background.
Osteology. Osteology based on five cleared and stained specimens. Vertebrae 11 precaudal and 16 caudal (total 27)(5 specimens); dorsal pterygiophore formula 3- 221110 (4) or 3-212110 (1); anal pterygiophores anterior to first haemal spine 2 (5); procurrent rays 6/ 6 (upper/lower) (2), 7/7 (2), 7/6 (1); segmented branched caudal rays 7/7 (2), 7/6 (3); segmented unbranched caudal rays 2/1 (2), 1/1 (2),?/1 (1); epurals 1 (5); hypurals 1/2 fused to hypurals 3/ 4 halfway (5).
Etymology. The specific name honors Alfredo Carvalho-Filho ─Alfie, as he calls himself, a self-made ichthyologist in his spare time, for his contribution to the advancement in the diversity and taxonomy of Brazilian marine fishes and his friendship.
Distribution, habitat and natural history. Gobiosoma alfiei is currently known only from Brazil, from Anchieta, Espírito Santo to Iparana, Ceará. Most specimens from Espírito Santo were collected in shallow (<0.5 m) tidepools within a flat reef. The tidepools are, generally, depressions in the calcareous material with a sand and gravel substrate, tidepool edges covered by algal turf, soft macro-algae, coralline algae and a few soft and hard corals. The reef surface is generally flat and pools frequently interconnect with each other through both overflows and an extensive system of dissolution holes and channels (see Joyeux et al. 2009 for illustration). Sympatric gobiid species, that share the same pools but not necessarily the same microhabitat, include Bathygobius geminatus Tornabene, Baldwin & Pezold 2010 (most abundant), Bathygobius soporator (Valenciennes 1837) , Barbulifer enigmaticus (Joyeux, Van Tassell & Macieira 2009) , Coryphopterus glaucofraenum Gill 1863 and Ctenogobius saepepallens (Gilbert & Randall 1968) (least abundant) (Joyeux et al. 2009; Macieira & Joyeux 2011). Gobiosoma alfiei is not limited to the intertidal zone. Underwater observations in the state of Alagoas (northeastern Brazil) at about three meters depth (low tide) by one of us (JCJ) showed several G. alfiei living on (and in) a rock outcrop located under a reef ledge. The outcrop was composed of encrusting coralline algae, interspersed with filamentous and turf-like algae. Mostly, isolated (single) G. alfiei were observed perched at the entrance of a small hole in the calcareous rock, occasionally biting at the substrate, and darting into the hole at the slightest disturbance. In one occasion, an individual was observed chasing another, both finally entering the same hole and not reappearing during the observation period. In the state of Paraíba (northeastern Brazil), the species was observed by one of us (JCJ) in subtidal areas (about three meters depth) under a small, flat, rounded stone lying on top of a flat clay substrate. Upon lifting the stone, the fish disappeared into a small round hole in the substrate. In a 15 cm deep tidepool in the state of Espírito Santo, one of us (JCJ) observed G. alfiei entering and exiting one of the numerous holes in the clay along the side of the pool. Swimming was limited to close contact with the bottom, in small (1–10 cm) darts, starting and ending in protected areas (i.e., under stones, near holes, into cavities). Breeding males were observed perched near holes for extended periods, moving up to 30 cm away, before entering/exiting the hole.
Conservation. Least Concern (Assessment done by MMA/ICMBio; Ministério do Meio Ambiente/ Instituto Chico Mendes de Conservação da Biodiversidade). No known specific threat, but the anthropogenic modifications on coastal marine ecosystems could be a threat in the near future.
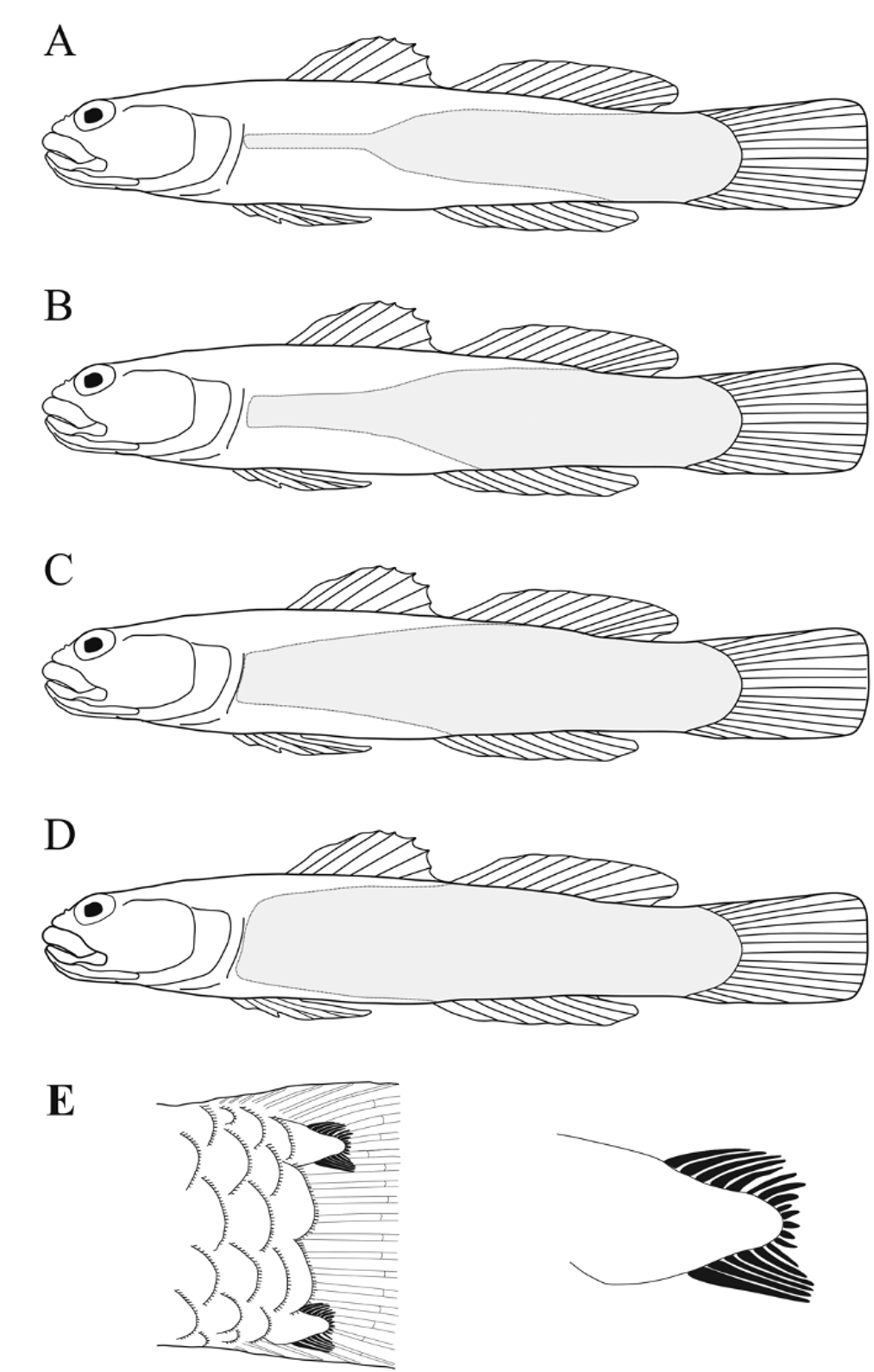


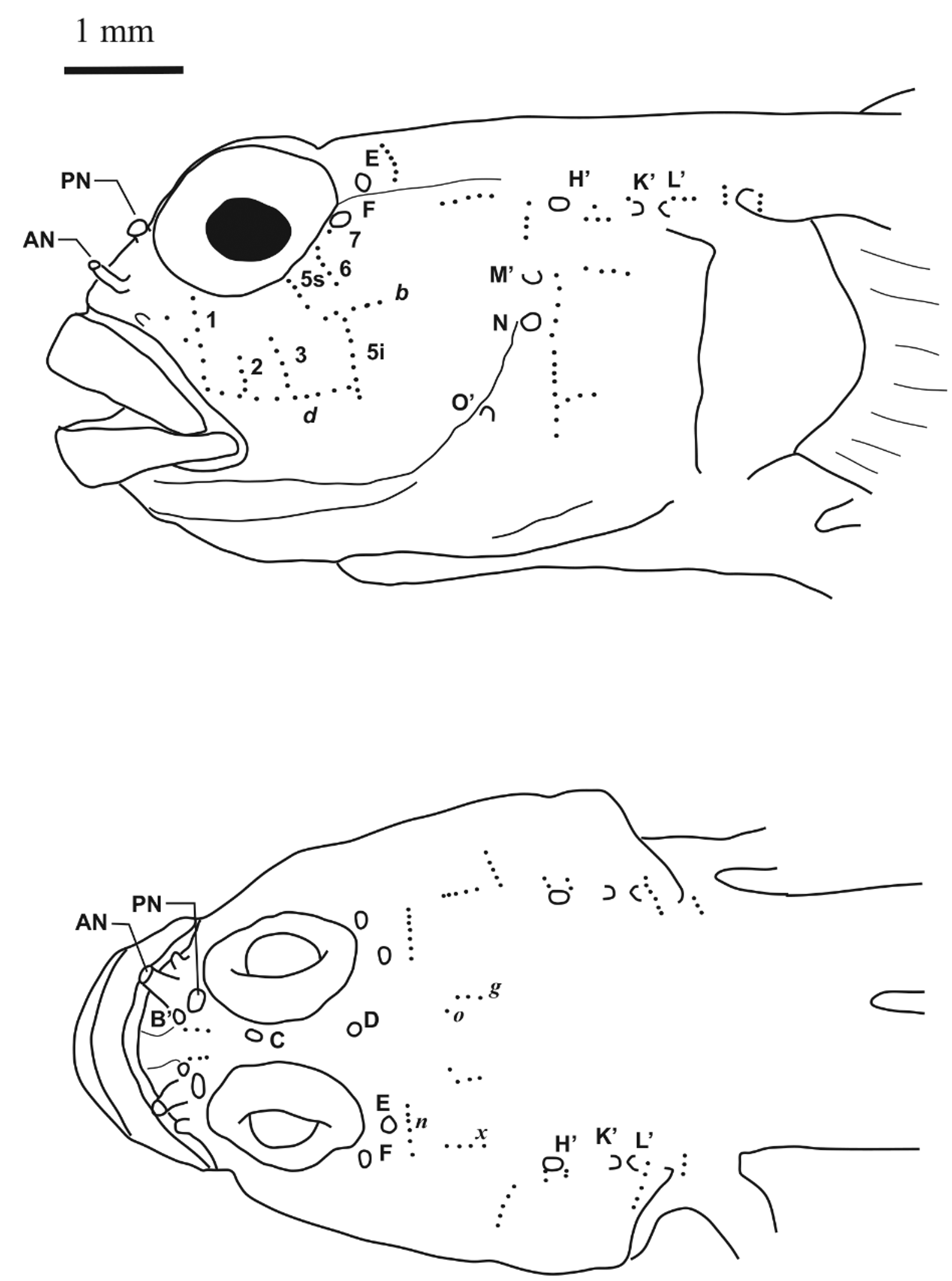
TABLE 4. Meristic and morphometric data for 52 examined specimens of Gobiosoma alfiei spec. nov. Morphometric data expressed as percent of standard length, of head length (†) or of jaw length (‡). n = number of individuals, including types.
| Meristic characters | Holotype | n | Range | Mode |
|---|---|---|---|---|
| First dorsal fin | VII | 45 | VI–VII | VII |
| Second dorsal fin | I,11 | 48 | I,8–I,11 | I,11 |
| Anal fin | I,9 | 47 | I,5-I,9 | I,9 |
| Pectoral fin - right | 18 | 42 | 17–20 | 18 |
| Pectoral fin - left | 18 | 39 | 17–20 | 18 |
| Pelvic fin rays | 5 | 48 | 5–5 | 5 |
| Segmented rays in caudal fin | 15 | 47 | 14–17 | 17 |
| Branched rays in caudal fin | 14 | 41 | 11–15 | 14 |
| Scales in longitudinal series | 29 | 51 | 3–33 | 29 |
| Morphometric characters | Holotype | n | Range | Mean ± SD |
| Total length (mm) | 20.6 | 52 | 9.2–27.8 | 20.5 ± 4.1 |
| Standard length (mm) | 17.2 | 52 | 8.4–22.8 | 16.8 ± 3.3 |
| Standard length (% TL) | 83.6 | 52 | 79.2–91.3 | 82.2 ± 1.8 |
| Head length | 29.1 | 48 | 27.1–37.6 | 29.9 ± 1.7 |
| Head depth | 18.6 | 48 | 17.9–24.4 | 20.3 ± 1.5 |
| Head width | 20.4 | 48 | 19.0–27.2 | 23.0 ± 1.8 |
| Postorbital length | 17.5 | 48 | 14.7–19.3 | 17.2 ± 0.9 |
| Upper jaw length | 5.8 | 44 | 4.0–9.6 | 6.4 ± 1.3 |
| Interorbital width | 1.2 | 44 | 0.6–2.1 | 1.4 ± 0.4 |
| Eye diameter | 8.1 | 48 | 6.7–9.6 | 8.1 ± 0.7 |
| Pupil diameter | 2.9 | 39 | 2.9–4.6 | 3.7 ± 0.4 |
| Anterodorsal length | 8.1 | 48 | 6.0–14.8 | 8.6 ± 1.5 |
| Face barbel length | 1.7 | 25 | 0.3–2.3 | 1.2 ± 0.5 |
| First dorsal spine length | 12.2 | 33 | 8.5–15.0 | 12.1 ± 1.5 |
| Depth of body at D1 origin | 17.5 | 46 | 15.2–23.4 | 18.9 ± 1.7 |
| Pectoral fin length - left | 25.0 | 35 | 16.5–28.6 | 24.2 ± 2.4 |
| Pectoral fin length - right | - | 37 | 21.6–28.3 | 24.7 ± 1.7 |
| Pelvic fin length | 17.5 | 48 | 13.9–23.9 | 18.6 ± 1.8 |
| Caudal peduncle depth | 13.4 | 44 | 12.2–15.4 | 13.8 ± 0.8 |
| Caudal fin length | 22.7 | 48 | 16.7–27.0 | 23.5 ± 1.9 |
| Postorbital length † | 60.0 | 48 | 45.5–64.2 | 57.5 ± 3.5 |
| Upper jaw length † | 20.0 | 44 | 12.8–32.7 | 21.5 ± 4.6 |
| Interorbital width † | 4.0 | 44 | 1.9–7.1 | 4.8 ± 1.4 |
| Eye diameter † | 28.0 | 48 | 21.8–33.3 | 27.1 ± 2.3 |
| Pupil diameter † | 10.0 | 39 | 10.0–16.7 | 12.5 ± 1.4 |
| Face barbel length † | 6.0 | 25 | 1.0–7.7 | 4.2 ± 1.6 |
| Eye diameter ‡ | 140.0 | 44 | 76.5–220.0 | 132.9 ± 31.5 |
| UFPB |
Departamento de Sistematica e Ecologia |
| AMNH |
American Museum of Natural History |
| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
| ZUEC |
Museu de Zoologia da Universidade Estadual de Campinas |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| SIO |
Scripps Institution of Oceanography |
| IPK |
Institut fuer Pflanzenschutzforschung |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Gobiosoma alfiei
| Van Tassell, James L., Joyeux, Jean-Christophe, Macieira, Raphael Mariano & Tornabene, Luke 2015 |
Gobiosoma parri
| Gomes 2014: 1001 |
Gobiosoma hemigymnum (
| Macieira 2011: 311 |
Gobiosoma hemigymnum
| Macieira 2011: 311 |
| Joyeux 2009: 64 |
Gobiosoma
| Joyeux 2004: 1281 |
Gobiosoma spilotum (
| Rocha 1998: 564 |