
Gypsonictops lewisi, Sahni, 1972
|
publication ID |
https://doi.org/ 10.5281/zenodo.3382461 |
|
DOI |
https://doi.org/10.5281/zenodo.4710540 |
|
persistent identifier |
https://treatment.plazi.org/id/1A7187CF-FFA3-1710-FE78-F65FEA495088 |
|
treatment provided by |
Plazi |
|
scientific name |
Gypsonictops lewisi |
| status |
sp. nov. |
Gypsonictops lewisi , new species
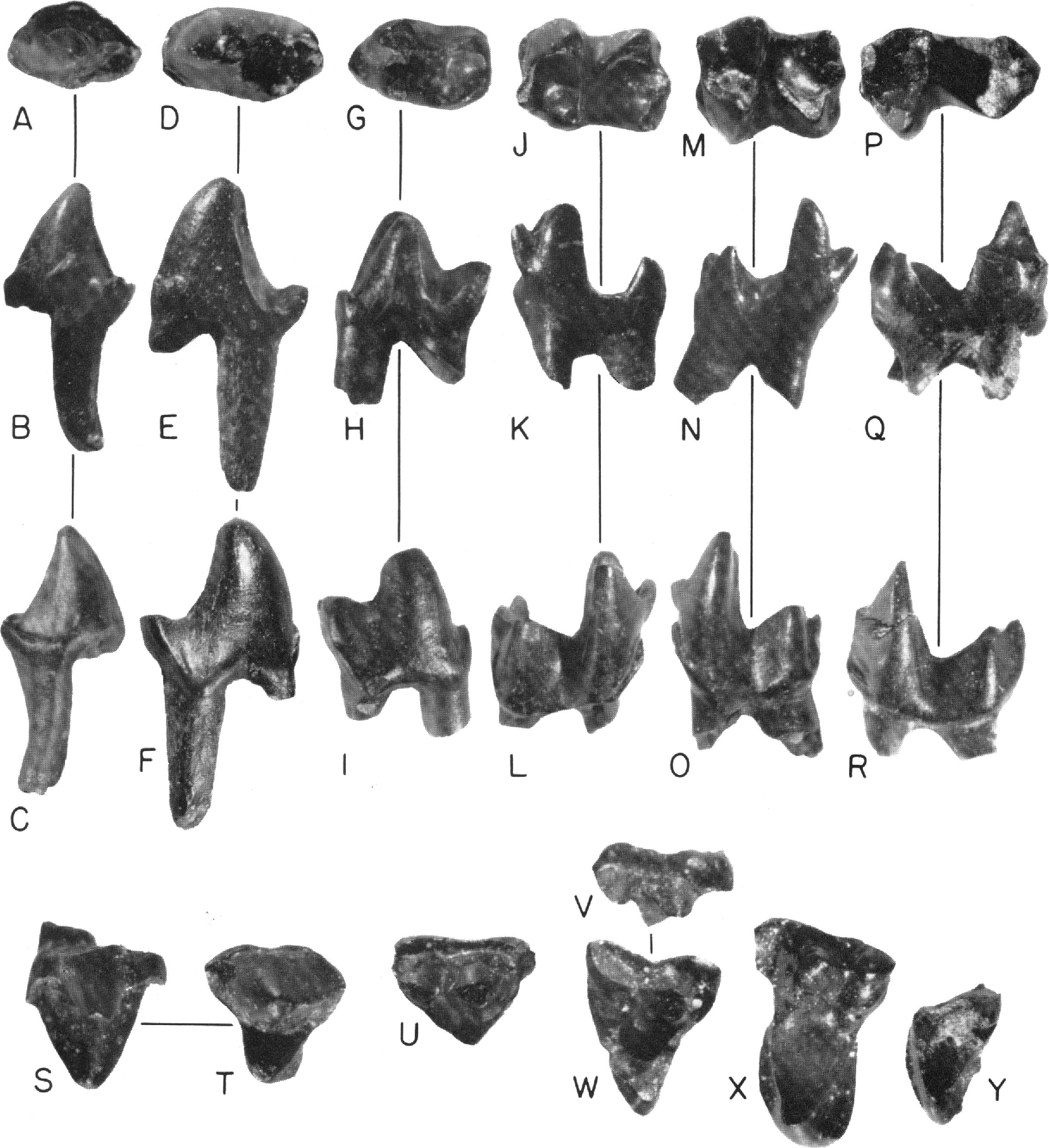
Figure 15 View FIG
ETYMOLOGY: For Captain Meriweather Lewis, pioneer explorer of the Missouri River.
HOLOTYPE: AMNH 77429, a right P4.
TYPE LOCALITY: Clambank Hollow, Chouteau County, Montana.
KNOWN DISTRIBUTION: Campanian, Judith River Formation.
DIAGNOSIS: Similar in most respects to Gypsonictops hypoconus and G. illuminatus , but with less molariform P3 and P2, and a more molariform P3.
DESCRIPTION: Lower dentition: P2; two teeth are probably P2s referable to G. lewisi . Their size and morphology sets them apart from the posterior premolars of a reconstructed dental series. Their resemblance to those of placental mammals is indicated by presence of a small metaconid that is not fully separated from the long and slender protoconid, and a shallow talonid bearing two cusps. Anterior lower premolars of Cretaceous metatherians have a protoconid that is linked to a posterior accessory cusp by a median ridge, and cingula that may be developed on either side of this ridge.
A double rooted tooth, AMNH 77426 (fig. 15A-C), is smaller than P3. The specimen lacks its anterolabial part and the anterior root. The protoconid is a large cusp with a steep anterior margin. A small metaconid arises from the lingual cingulum and is not well separated from the protoconid. Buccal to the protoconid there is a well-developed labial cingulum that terminates at the labial accessory cusp of the talonid. This latter cusp is also linked to the apex of the protoconid by a shear ridge. There is at least one other cusp lingual to, and higher than, the labial accessory cusp. The talonid basin is shallow.
The identification of the tooth as a P2 is based primarily on occlusal relationships and comparison with P3. Although present, the talonid is too shallow to be functional and probably occluded with a p2 which had a small protocone without root support. Unfortunately, p2 of Gypsonictops lewisi has not been found. P2 of G. lewisi is smaller than P3, and it lacks the lingual ridge arising from the posterior side of the protoconid apex.
P3; reference to specimen AMNH 77428 as P3 is based in part on the occlusal relationship of the tooth and in part on the development and degree of separation of the major cusps. This submolariform tooth has an anterior cingulum extending from the anterolingual side of the protoconid ventrally to the anterolabial side (fig. 1 5D-F). A small cuspule, more a basal swelling than a distinct cusp, is present at the paraconid position, and probably represents that cusp. The most proininent cusp is the protoconid, which is high and conical with convex anterior slopes. Two ridges arise from the posterior border of the protoconid; one extends from its lingual slope to the heel of the tooth, and the other extends more medially and joins the two major cusps of the talonid. A groove is present on the posterior wall of the protoconid between the two ridges. The metaconid is not clearly visible, represented only by a swelling on the lingual ridge at a point one-half the height of the protoconid.
The talonid is basined and deeper than that of the supposed P2. Two cusps are situated close together and probably correspond to the hypoconid-hypoconulid. A ridge bearing a small cuspule arises from the more lingual of these cusps and probably represents the entoconid. A shallow cingulum marks the posteroventral border of the tooth on the labial side.
P3 of Gypsonictops lewisi is distinct from P3 of G. petersoni , G. illuminatus , and G. hypoconus . The Campanian specimen has a better developed and more basined talonid which bears two cusps. Simpson (1951, pp. 10, 17) commented on the P3 "being simple and abruptly different from the molariform P4."
P4; three teeth can be considered P4s on the basis of their similarity to similarly placed teeth in jaws of Gypsonictops hypoconus from the Lance and Hell Creek formations, CM 11657, and UMVP 2316, 2317, and 2311. A distinct paraconid is present on the anterobasal cingulum in front of the groove separating the metaconid from the protoconid in AMNH 77429 (fig. 15G-I). Metaconid is not fully separated from the protoconid and in all three specimens at hand, it is a slender, conical cusp that is much smaller than the protoconid. A high ridge links the posterior wall of the protoconid to the hypoconid. An anterior ridge arises from the protoconid apex, and joins the paraconid ventrally in AMNH 77429. This is similar to the condition seen in the P4, UMVP 2311.
The talonid of P4 is basined and well developed, and all three cusps are represented. The hypoconulid is separated from the hypoconid, and it is only slightly smaller than the hypoconid. The entoconid is a feebly developed cusp situated on the lingual ridge of the talonid anterior to the other cusps. Labial to the crista obliqua, the crown extends ventrally, and a distinct cingulum is present on the posteroventral border.
P4 of G. lewisi is distinguishable from that of other species because it has a stouter, broaderbased protoconid, it has less separation of the metaconid from the protoconid, and it has a less-developed talonid.
M1 and M2; there are five lower molars that are either M1 or M2. The criterion used here to distinguish between molars is the tendency for the talonid to get progressively narrower (transversely) toward M3. On this basis, two teeth are probably M1s and the remainder M2s. One tooth, AMNH 77432 (fig. 16J-L), is tentatively identified as M1, and follows the basic therian pattern. Its trigonid is anteroposteriorly compressed, and its smallest cusp is the anteriorly projecting paraconid, the worn apex of which is only slightly labial to the apex of the metaconid. Its paraconid is connected to the protoconid by a ridge. A short anterobasal cingulum is present labial to the paraconid. The metaconid is a well-developed cusp and slightly smaller than the protoconid. The talonid of AMNH 77432 is basined and wide, and the hypoconid is large with a short cingulum extending down its posterior wall. The hypoconulid is well separated from the adjacent talonid cusp and is medial in position. The entoconid appears to be the highest talonid cusp. Of the two roots, at the enamel base, the anterior one is more anteroposteriorly compressed.
M2; specimen AMNH 77434 (fig. 15M-O) is probably an M2. It differs from M1 by having a shorter, more anteroposteriorly compressed, and narrower talonid.
M3; a possible M3 is specimen AMNH 77438 (fig. 15P-R) because its talonid is elongated anteroposteriorly and is much narrower in comparison to the trigonid width of either M1 or M2. The anterobasal cingulum flattens to form a small shelf. The trigonid is similar to those of the other molars but the talonid is very different. It narrows considerably at the point where the crista obliqua intersects the protolophid (metaconid-protoconid ridge). The three cusps of the elongate talonid are well developed, the hypoconulid and the entoconid are situated closer to each other.
UPPER DENTITION: A reconstruction of the upper dentition of Gypsonictops was made by Simpson (1951) and had been accepted until recently when doubts about the identity of P4 were expressed by McKenna (1960) and Slaughter (1968a). According to these authors, P4 figured by Simpson (1951) is most probably P3. P4, then, is probably an even more molariform tooth. McKenna based his conclusions on a fragmentary maxilla with p2, p3, and roots of P4, UCMP 51386. This specimen is currently under study by W. A. Clemens in his survey of the eutherians of the Lance Formation. The holotype maxilla of Gypsonictops illuminatus Lillegraven (1969) , UAPC 2447, with P3 to M3, from the upper part of the Edmonton Formation, conclusively demonstrates that McKenna (1960) and Slaughter (1968a) are correct in their interpretation. Gypsonictops material from the Judith River Formation, although clearly ancestral to the Maestrichtian species, differs from them in structure of its premolars. Premolars of G. lewisi are less molariform than those of later species.
P3; there are seven teeth in the collection that I consider to be P3 rather than p 2 because of the occlusal relationship with the submolariform lower anterior premolars, particularly P2, and because it is unlike p2 in the Gypsonictops maxilla, UCMP 51386.
Specimen AMNH 77442 (fig. 15S, T) is subtriangular in shape. Its most prominent cusp is the large, conical, broad-based paracone. Labial slopes of the paracone descend into a welldeveloped ectocingulum particularly in the parastylar and metastylar regions. The stylar shelf narrows at the ectoflexus where the paracone forms the buccal edge of the crown. A sharp crest, the metacrista, extends posteriorly from the apex of the paracone to the metastyle. Just anterior to the metastyle a slight swelling is present on the stylar shelf. The protocone is much smaller than the paracone with steeper lingual slopes. A definite postcingulum is present in three of the seven complete teeth and in two of these three teeth a smaller anterior cingulum is also present. Two ridges link the protocone to the paracone, but conules are absent. These three teeth have roots with rounded cross sections, and the most lingual root is placed beneath the protocone.
P3 is submolariform because the paracone is undivided and has a sharp metacrista, and because the protocone has root support and lingual cingula. P3s of the later species of Gypsonictops are more molariform, chiefly because the metacone is more separated from the paracone. Separation of the metaconid on the lingual side of the paraconid is seen in a specimen of Gypsonictops , UMVP 2316, from the Hell Creek beds. P3 of G. lewisi is approximately at the same stage of molarization as P3.
P4; there is only one tooth, AMNH 77451 (fig. 15U), that is considered P4. The reasons for this are not its similarity to P4 of G. hypoconus , as identified by Simpson (1951) (now considered to be P3), but because of its degree of molarization relative to P3 and Ml, and because of its occlusal relationship to P4. This specimen, AMNH 77451, lacks the area lingual to the paracone and metacone. A trace of a paraconule was present but during preparation this region broke away and thus the protoconule is not visible in the illustration. The tooth is more molariform than P3, as indicated by the greater separation of the paracone from the metacone. These cusps, however, are undivided at the base and only a narrow valley separates their apexes. Absence of a prominent ectoflexus and less transverseness also imply that it is less molariform than the upper molars. A small protocone with root support, a paraconule, and both precingulum and postcingulum must have been originally present. P4 of G. lewisi closely resembles P3 of G. hypoconus . The parastylar area is more prominent than the metastylar area and no ectoflexus is present. The apex of the metacone is separated from the larger paracone by a short centrocrista and is connected to the metastyle by a metacrista.
M1; three teeth have been identified as Mls. They are more molariform than the supposed P4, less transverse, with less prominent lingual cingula than M2. The stylar shelf extends above the entire labial margin of the crown in AMNH 77452 (fig. 15V, W), and the parastylar area is well developed with a prominent parastyle connected to the paracone by a short, low ridge. The metastyle is joined to the metacone by a metacrista. The paracone is taller than the metacone and separated from it by a deep valley. Conules are present, the preparaconular wing of the paraconule is bounded anteriorly by a ridge extending anterobuccally, and a prominent premetaconular wing is also present. The protocone is a narrow, thin cusp with nearly vertical lingual slopes. The preprotocrista and the postprotocrista join the paraconule and the metaconule, respectively. The postcingulum is larger than the precingulum and bears a worn area representing the hypocone.
M1 of G. lewisi is similar to the upper molar, USNM 5044, figured by Simpson (1951, fig. 2b).
M2; there is only one worn but complete M2 (AMNH 77456, fig. 15X). It is distinguished from M1 by having a narrower stylar shelf with a more anterobucally expanded parastylar region, by being shorter and more transverse, and by fuller development of the precingulum and the postcingulum.
The paracone and metacone are set buccally, and the precingulum is better developed than on M1 and extends from a point just ventral to the paraconule, to the anterolingual side of the protocone. The postcingulum, bearing a hypocone, is even larger than the precingulum, extending from a point ventral to the base of the metacone, and terminating on the posterolingual side of the protocone. The tooth is three rooted, and its lingual root is very compressed anteroposteriorly. Its labial roots are small in comparison and have rounded cross sections.
M3; the third upper molar is known by a lingual fragment, AMNH 77457 (fig. 15Y). Its paracone is missing, but the metacone is a small, conical cusp. The paraconule is better developed than the metaconule, and the postcingulinm is broken in the specimen.
DISCUSSION: Patterson's (1956) description of mammals at the eutherian-metatherian grade has greatly increased our knowledge about placental origins. Mammal remains recovered from the Trinity Sandstone of Texas, of Albian age (Lower Cretaceous), indicate that they had already differentiated into placental and marsupial types, as based on presence or absence of submolariform premolars. Submolariform premolars indicate a specialization unknown in marsupials ( Slaughter, 1968a, 1968 b, 1968c). The next record of eutherians in North America is during the Campanian, and the new material from the Judith River Formation is the principal source of this information. During the Maestrichtian, placentals increased in diversity in various western states of North America, and at the close of the Cretaceous represent the orders: Insectivora, Deltatheriidia (sensu Van Valen, 1966), Condylarthra, and Primates (Sloan and Van Valen, 1965; Van Valen and Sloan, 1965).
Gypsonictops was first described by Simpson (1927a) on the basis of isolated upper teeth from the Lance Formation. These teeth had originally been assigned by Marsh (1892c) to Batodon and Telacodon . Simpson (1951) brought together previous information on this Cretaceous insectivore and made a hypothetical reconstruction of its dentition. This reconstruction was based partly on upper teeth assigned to Gypsonictops and partly on lower teeth assigned to Euangelistes . Later, it was found that probably two different species were involved, because the dimensions of the lower teeth were considerably larger than those of the upper teeth. Two species were thus retained, G. hypoconus , based on the smaller upper molars, and G. petersoni , based on the larger lower molars.
Although the hazards of such a hypothetical reconstruction were obvious to Simpson (1951), subsequent finds have shown that, at least in the association of upper and lower dentitions, he was correct, but owing to misidentification of P3 as P4 and of P4 as an upper molar Simpson was led to believe that Gypsonictops possessed simple, not incipiently molariform, pl_p3.
The placental mammal, G. lewisi , from the Judith River Formation, is represented by about 45 teeth, comprising slightly over 5 per cent of the total mammal teeth recovered, and a calcaneum quite similar to that of Prodiacodon ( Szalay, 1966) . It probably belongs to Gypsonictops and is currently under study by W. Nelson. The majority of the isolated teeth are broken or worn. None of the edentulousjaws can be referred with any certainty to the species. Still, by comparison with other eutherian material in the Maestrichtian of Alberta and Montana, it has been possible to identify the series of teeth, P2 to M3, and P3 to M3. Teeth anterior to P2 and P3 may be present in the collection, but are as yet unidentified. If the reconstruction of the dentition of G. lewisi proves to be correct, then the Campanian species differs from those of the Maestrichtian, at least at the species level.
Absence of Cimolestes and other members of the Palaeoryctidae from the Judith River Formation lends some credence to the viewpoint that deltatheridians were an immigrant group that invaded the coastal plains east of the rising Rocky Mountains sometime during the Maestrichtian. Gypsonictops is considered here to be a primitive placental and, by virtue of its presence in pre-Maestrichtian sediments, is a likely ancestor for some later North American eutherians.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.