
Heteromeringia nigrifrons KE, 1903
|
publication ID |
https://doi.org/ 10.21248/contrib.entomol.57.1.37-80 |
|
DOI |
https://doi.org/10.5281/zenodo.4793477 |
|
persistent identifier |
https://treatment.plazi.org/id/763A87EE-FFD2-FF8A-FEBD-FF72FC127E4D |
|
treatment provided by |
Carolina |
|
scientific name |
Heteromeringia nigrifrons KE, 1903 |
| status |
|
Heteromeringia nigrifrons KE, 1903 View in CoL
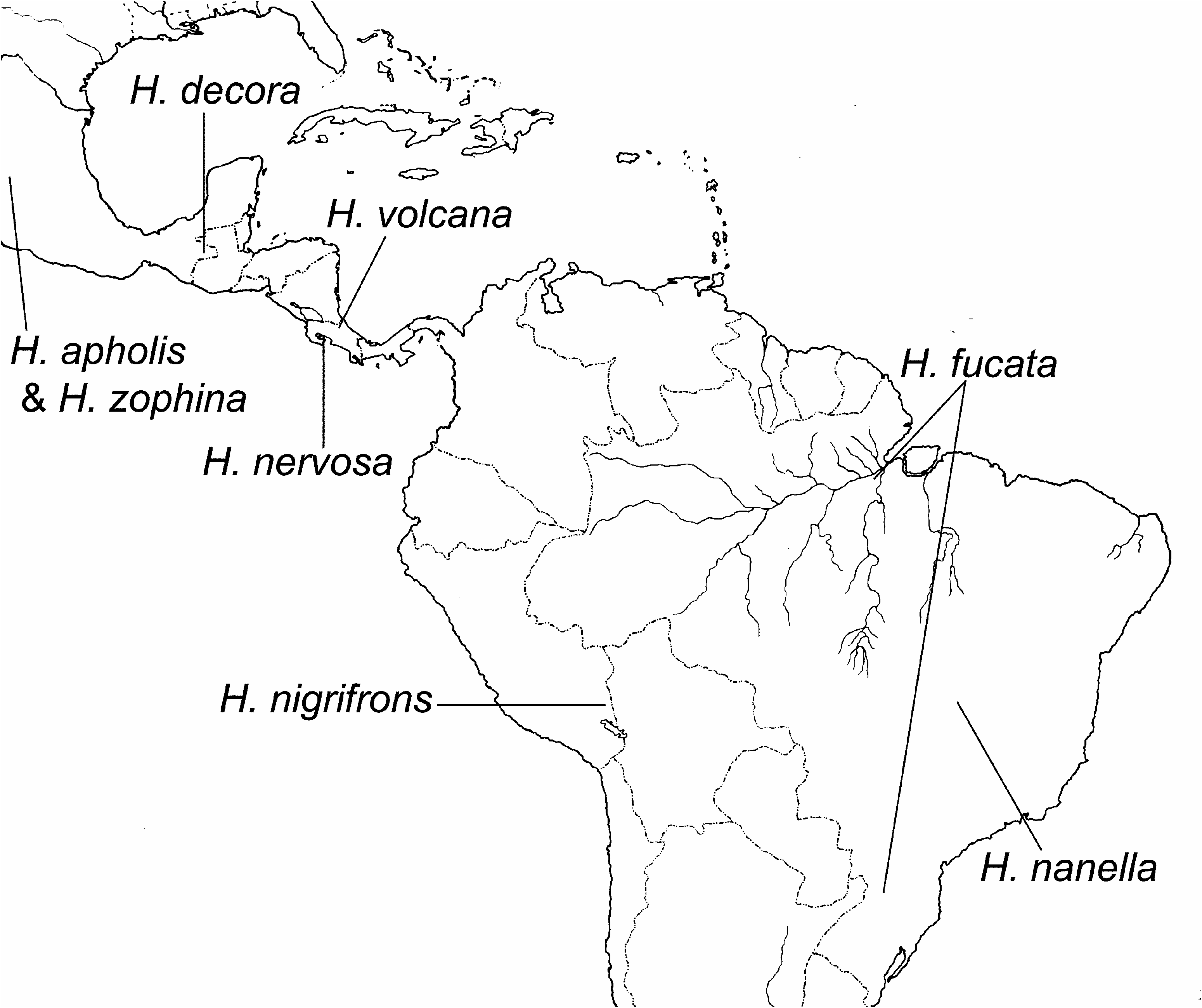
( Map 3 View Map 3 )
Heteromeringia nigrifronsnigrifrons KERTESZ, 1903: 568 . MELANDER & ARGO, 1924: 30.
Description [translated from KERTESZ (1903)]
Female
Black, with weak luster. Frons, face, gena and occiput black. Orbits from base of antenna to slightly beyond midpoint of frons narrowly brownish yellow. Antenna pale brownish yellow; third segment more or less reniform and with black apex; arista black, pubescent. Proboscis and palpus black, the latter reddish yellow in apical half. Bristles of head black. Genae ca. 1/6 as high as head. Vibrissa moderately long, behind it only very short setulae. Thoracic dorsum dull black, with appressed very short and weak ferruginous pubescence; bristles black. Pleura shining black. Legs reddish yellow. Front coxae pale yellow. Front femur with exception of basal half and knee, mid femur toward (i.e., excluding) apical fourth, and tibiae excluding knees yellow. Mid and hind tibiae with black [? ventral] apical spur. Femora with short bristles on ventral surface (bristles of hind femur longest). Front tibia and tarsi with short black setulae. Wing, excluding yellowish basis and hind margin, with smoky tinge; infuscation strongest at front margin. Third and fourth longitudinal veins parallel, last sector of fourth vein five times as long as penultimate sector. Halter yellowish white. Length: 4.3 mm; wing length: 3 mm.
Female terminalia
Not dissected.
Male
Unknown.
Holotype: PERU. Vilcanota (1 , HNHM).
Comments
The holotypes of Heteromeringia nigrifrons and H. flavifrons were examined on our behalf by Dr. L. Papp in Hungary, allowing us to add the following notes to the above description: there is no upcurved bristle on posterodorsal surface of the anepisternum; the gena and occiput are shiny; the postvertical bristle is slightly shorter than the ocellar bristle; there are two pairs of dorsocentral bristles; the M 1+2 ratio is 6.9. Heteromeringia nigrifrons is the only New World Heteromeringia outside of the H. czernyi group with yellow fore tibiae.
Recombinations
Two species previously treated as Heteromeringia are no longer included in the genus. Heteromeringia annulipes JOHNSON, 1913 , described from North Carolina, was recently moved to Craspedochaeta CZERNY, 1903 as a senior synonym of the predominantly neotropical C. piceoflava SÓOS, 1962 ( LONSDALE & MARSHALL, 2006b). The Peruvian H. dimidiata HENNIG, 1938 (holotype label data: Peru. Ucayalifluss, Unini, 20.x.1903, Schnuse (1 , SMTD)) belongs to a clade of neotropical SobarocephalaSobarocephala including S. liturata MELANDER & ARGO, 1924 and is therefore treated here as Sobarocephala dimidiata (HENNIG, 1938) comb. n. This species was mistakenly treated as HeteromeringiaHeteromeringia because its dark colouration gave it a resemblance to H. fumipennis MELANDER & ARGO, 1924 (HENNIG, 1938).
Replacement names
A replacement name is here provided for the South African/ Seychelles species Heteromeringia nigrifrons LAMB LAMB, 1914 , a primary junior homonym of the neotropical H. nigrifrons KERTESZ, 1903 . The replacement name, H. tephrinos nomen nov., is Greek for “ash-coloured”, in reference to the infuscated halter.
Relationships
Most New World Heteromeringia can be placed into one of two relatively common species groups: the widespread H. nitida group and the entirely Neotropical H. czernyi group. Three Neotropical species ( H. apholis , H. decora and H. nigrifrons )), and several species from Europe, Southeast Asia and Fiji, cannot be placed in either of the above species groups.
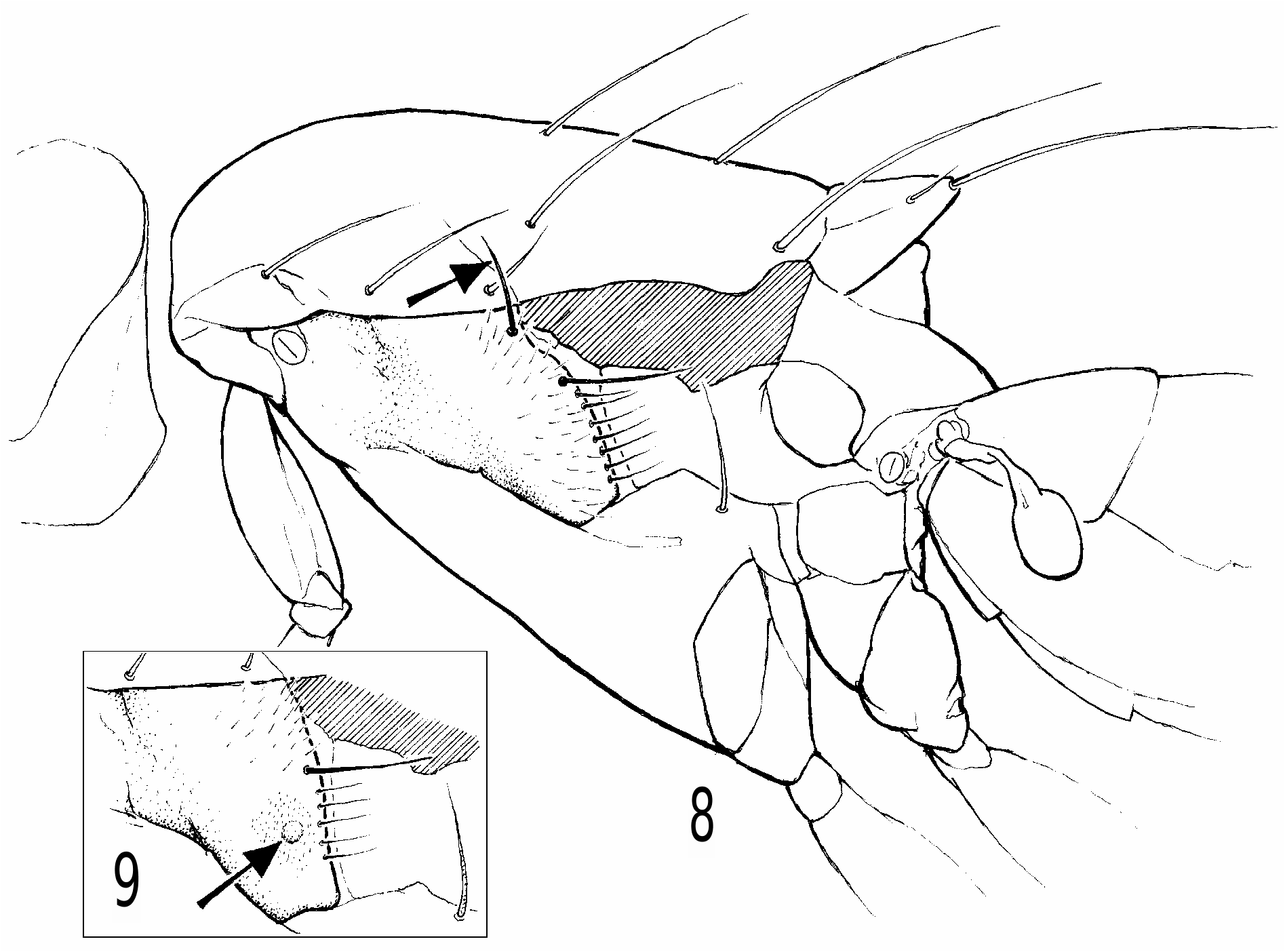
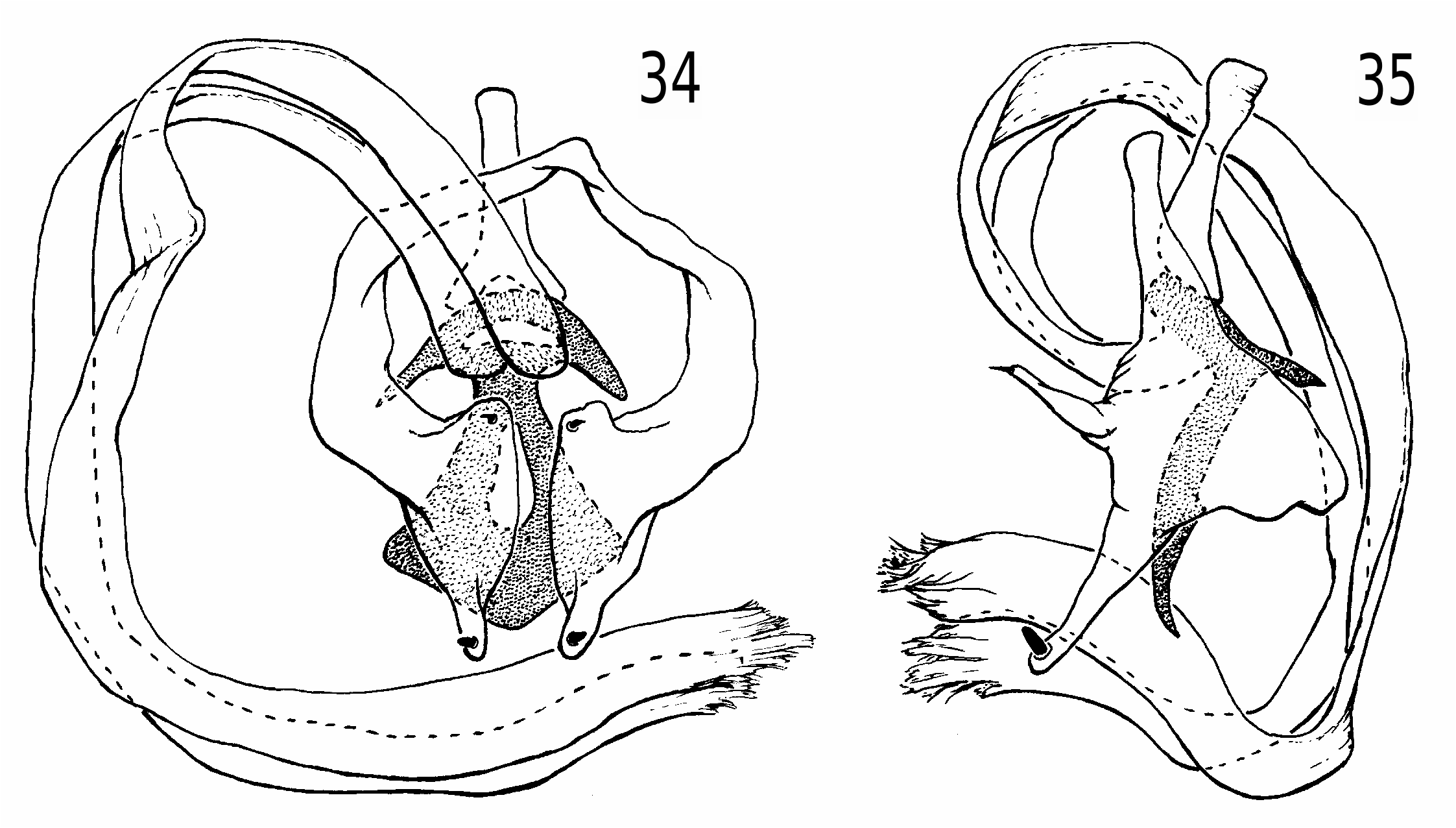
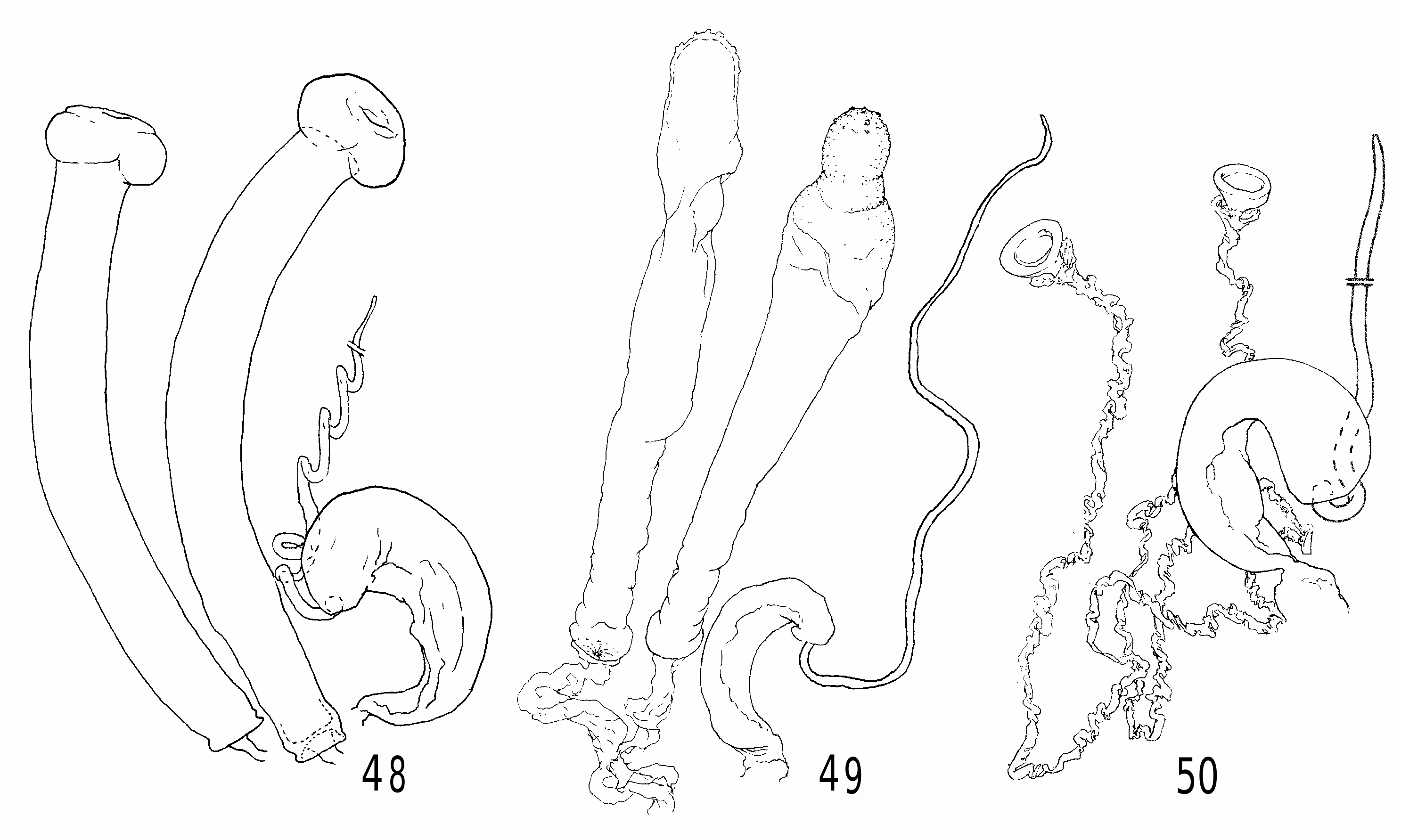
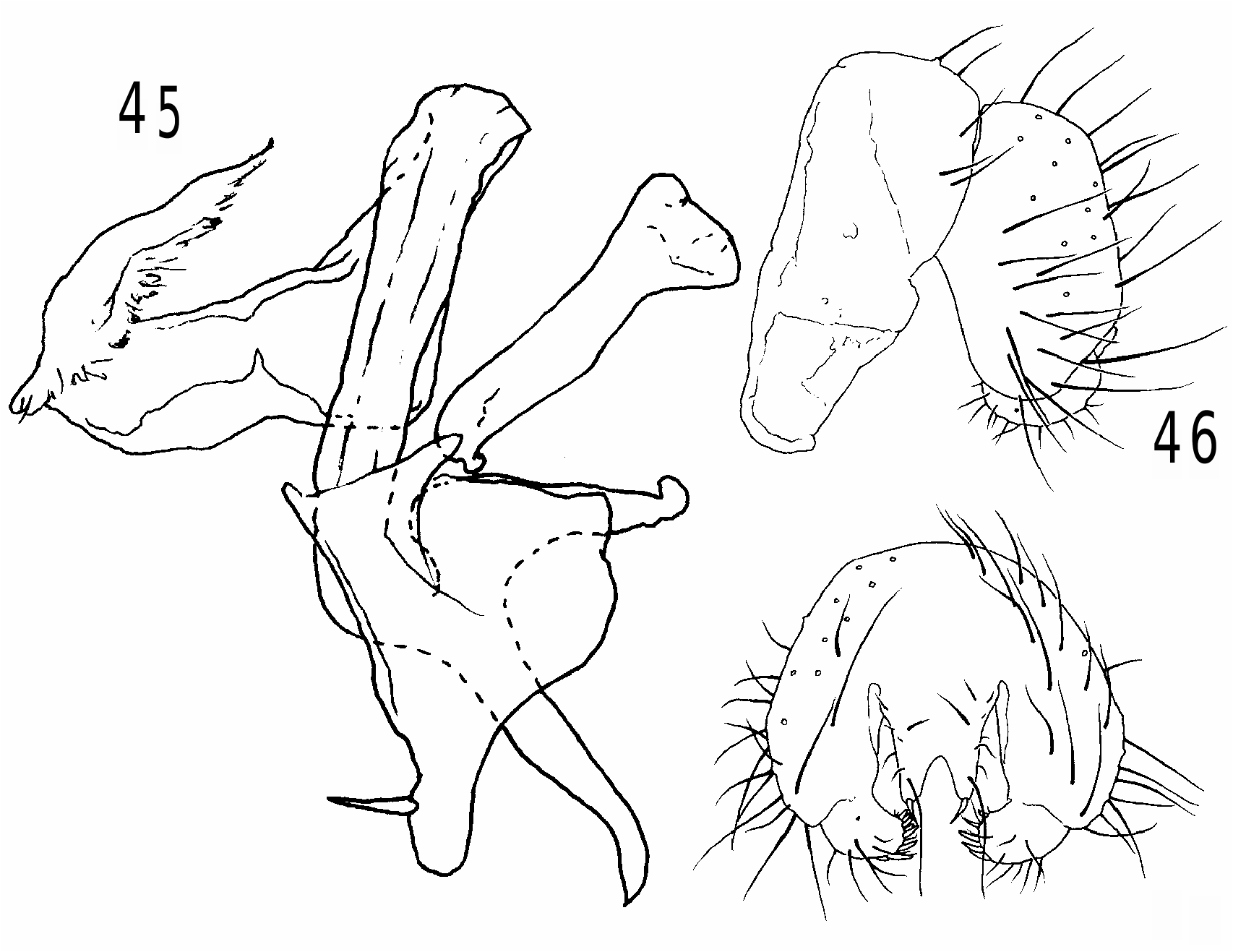
Heteromeringia czernyi species group. The H. czernyi group contains H. czernyi , H. flavipes , H. aphotisma , H. flavifrons , H. lateralis , H. quadrisetaquadriseta and H. mediana , and is defined by the following characters: anterior genal bristle vibrissa-like; anepisternum with additional upcurved bristle in posterodorsal corner ( Fig. 8 View Fig ) (only retained in H. czernyi , H. flavipes and H. aphotisma ); arista loosely pubescent or short-plumose; pregonite with rounded distal nub ( Figs 34 and 35 View Figs 34-35 ); basiphallus with diamond-shaped to (somewhat) rectangular posteroventral plate behind point of attachment to distiphallus; phallapodeme never longer than hypandrium + pregonite; ventral receptacle large and wedge-shaped ( Figs 48-50 View Figs 48-50 ); spermatheca not telescoped.
Heteromeringia flavipesflavipes and H. czernyi are basal in the group, characterized by brown bristles, one distal and one medial cloud on the wing, hand-shaped scales on the surstylus ( Fig. 33 View Figs 31-33 ) (absent in some specimens), a prominent process on the inner face of the surstylus, and an elongate sac-like spermatheca with distal sculpturing ( Figs 48 and 49 View Figs 48-50 ). Both species extend into Florida, but H. flavipes is not found south of Costa Rica, and H. czernyiczernyi is widespread in both Central and South America all the way to Tierra del Fuego, which is by far the most southern record of a clusiid in the New World. The remaining six species in the group form a clade defined by four fronto-orbital bristles and a dark infuscation from the costa to the midpoint of the second radial cell (reduced in H. quadriseta ). Of these, H. aphotisma (southern Brazil) is the most ancestral because it retains dark colouration and an additional upcurved bristle in the posterodorsal corner of the anepisternum. The other four species are pale flies known either from Costa Rica ( H. lateralis )) or South America ( H. quadriseta : Ecuador, Peru; H. mediana : Brazil)), although the widespread Heteromeringia flavifrons has been collected in Bolivia, Brazil, Costa Rica, Ecuador and Honduras. The two exclusively South American species form a clade characterized by small inset surstyli ( Figs 44 View Figs 42-44 and 47 View Figs 45-47 ), an enlarged annulus ( Figs 43 View Figs 42-44 and 46 View Figs 45-47 ), and cerci that are thin and widely incised with one pair of relatively short, stout apical bristles ( Figs 44 View Figs 42-44 and 47 View Figs 45-47 ).
Heteromeringia nitida species group. The (single) defining synapomorphy of this widespread and species-rich group is the small, pale, disc-like structure on the male anepisternum (Fig. 9). This unusual character is unlikely to be subject to homoplasy and is an easily recognized synapomorphy. Many Heteromeringia belong to this group, but a World revision will be necessary to resolve its zoogeography and internal phylogeny, as well as the relationships between it and the remaining Heteromeringia .
Biology
Label data for New World specimens (mostly those we have collected in the Neotropics) suggest that some species of Heteromeringia aggregate around relatively open areas such as tree falls, forest borders, pastures and landslides. MCAC LPINE (1960) also notes that while most Australian clusiids were taken in rainforests along creeks, Heteromeringia species were collected in more open forests. We have collected H. fumipennis , H. czernyi , H. nitidanitida and H. flavifronsflavifrons with dung baits and dung traps. Heteromeringia nitida has also been collected using mushroom traps and has been observed on myxomycete sporocarps as described below. Heteromeringia flavifrons and H. czernyi have been collected from carrion. In Europe, H. nigrimana is known to be associated with dead wood in deciduous or mixed forests, predominantly on decaying logs ( ROHÁČEK OH, 1995). Australian specimens of H. norrisi MCAC LPINE, 1960 have been bred from rotting wood and one female was collected “in [a] log with termites” ( MCAC LPINE, 1960).
Agonistic behaviour
Like other clusiodine genera, including Craspedochaeta ( LONSDALE & MARSHALL, 2006b) , Clusiodes COQUILLETT, 1904 and Hendelia CZERNY, 1903 ( MARSHALL, 2000; MCAC LPINE, 1976; ROHÁČEK OH, 1995), Heteromeringia males engage in agonistic interactions at lek sites on fallen trees. One of us (SM) observed H. nitidanitida males visiting rotting oak logs in South Carolina over periods of several spring days in each of two consecutive years. Males appeared daily on bare patches of logs partially covered with myxomycete sporocarps, where each male seemed to patrol a territory of about six square cm. When two males appeared in the same territory, they folded their front legs back at the tibial-femoral joint before “boxing” with their elbows ( Fig. 1 View Fig ). Females were observed going in and out of beetle burrows on the same logs, and although pairs were seen in copula on the lek sites, no premating behaviour was noted. ROHÁČEK OH (1995) speculated that a recently fallen lime-tree log was used as a mating place for H. nigrimana , as seven of the nine specimens he collected off of this log were males. Males of H. pulla have been observed moving their wings in a similar fashion to sepsids, platystomatids and tephritids on the fallen bark and limbs of Eucalyptus trees, where they were also observed copulating with females ( MCAC LPINE, 1960).



| HNHM |
Hungarian Natural History Museum (Termeszettudomanyi Muzeum) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Heteromeringia nigrifrons KE, 1903
| Lonsdale, Owen & Marshall, Stephen A. 2007 |
Heteromeringia nigrifronsnigrifrons KERTESZ, 1903: 568
| ARGO, N. G. 1924: 30 |
| KERTESZ, K. 1903: 568 |