
Lobulia elegans (Boulenger)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4779.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:E7275795-8B03-4BE6-A25C-008D3325BE67 |
|
DOI |
https://doi.org/10.5281/zenodo.3851217 |
|
persistent identifier |
https://treatment.plazi.org/id/C65587E0-9374-FFEA-0F98-FF4AFE94FEC8 |
|
treatment provided by |
Plazi |
|
scientific name |
Lobulia elegans (Boulenger) |
| status |
|
Figures 1 View FIGURE 1 A–D, 2A, B, 3A
Lygosoma elegans Boulenger, 1897: 8 .
Type locality: Mt. Victoria , Papua New Guinea .
Lygosoma elegantoides Ahl, 1925: 20 (replacement name for Lygosoma elegans )
Lygosoma (Leiolopisma) elegans, Smith 1937: 224 .
Lygosoma (Leiolopisma) elegantoides lobulus, Loveridege 1945: 49 .
Scincella elegantoides, Mittleman 1952: 24 .
Lobulia elegans Greer, 1974: 9 .
Lobulia elegantoides Shea & Michels, 2008: 738 .
Material examined for rediagnosis. Papua New Guinea: Central Province: Mt. Victoria ( BMNH 96.10 .31.9, holotype), Mondo , 09.25ºS, 147.41ºE, 700 m a.s.l. ( BMNH 1935.5.10.143), Fane, 8.5530ºS, 147.0833ºE, 1400–1440 m a.s.l. ( BPBM 18691–93 View Materials ) GoogleMaps , NW slope Mt. Obree , 9.4456ºS, 147.9982ºE, 1330–1450 m a.s.l. ( BPBM 19594 View Materials ) GoogleMaps .
Diagnosis. A moderately sized species of Lobulia , adult SVL 59–66.5 mm; with a dorso-ventrally flattened body, distinct frontoparietals, two supralabials posterior to subocular, scales of temporal region not highly fragmented, lobules present on anterior margin of ear opening, subdigital lamellae 22–28 under 4th toe, mid-body scale rows 30–32, paravertebral scales 52–54, mid-dorsum with two rows of large dark-brown spots, top of tail base with one row of large dark-brown spots, sides brown with rows of small dark-brown spots, distinct pale dorsolateral stripe absent, light field on abdomen that lacks brown spotting ten scales wide, and under thighs and precloacal region white, without brown spotting.
Lobulia elegans can be distinguished from the other five species currently placed in the genus as follows: L. elegans differs from L. brongersmai in having separate (vs. fused) frontoparietals; from L. alpina Greer, Allison & Cogger and L. subalpina Greer, Allison & Cogger in having two (vs. three) supralabials posterior to the subocular supralabial and lacking (vs. having) a pale dorsolateral stripe; and from L. glacialis Greer, Allison & Cogger and L. stellaris Greer, Allison & Cogger in having a relatively unfragmented (vs. fragmented) temporal region, large chin shields abutting the infralabials (vs. separated by genials), and 22–28 (vs. 13–17) subdigital lamellae.
Description. This description is limited to six specimens from four localities in the central Owen Stanley Mts. of Central Province, Papua New Guinea, all of which are within 100 km of the type locality. All specimens I have seen from elsewhere but assigned to this species in museum collections represent other species that are typically undescribed; some of them are morphologically similar to L. elegans but genetically highly divergent (Slavenko et al. in prep.).
Rostral broad but shallow, approximately twice as wide as deep, projecting moderately onto tip of snout, its suture with frontonasal straight to shallowly concave; nasal elongate, widest anteriorly, usually entire but may have shallow dividing suture posteriorly (n = 2 of 12) or ventrally (n = 2 of 12); nasals widely separated, projecting approximately one-third of way onto dorsum of snout; nostril subcircular, centered within nasal; frontonasal large, pentagonal, extending laterally to dorsal level of nares, barely touching frontal; prefrontals moderate in size, narrowly separated, bordered below by two loreals; supraoculars four, anterior two contacting frontal; frontal kite shaped; one pair of frontoparietals in medial contact, in contact with posterior three supraoculars and in narrow contact with frontal ( Fig. 1A, C View FIGURE 1 ); interparietal smaller, kite-shaped; parietal eye spot absent; parietals in contact behind interparietal, in contact anteriorly with last supraocular, last supraciliary, and uppermost postsubocular; transversely enlarged nuchal on each side, larger than the scale separating it from the upper secondary temporal.
Anterior loreal smaller than posterior, higher than long; posterior loreal longer than high ( Fig. 1B, D View FIGURE 1 ); preocular longer than tall; presuboculars two (n = 11) or three (n = 1); postsuboculars five (n = 10) or six (n = 2), lowest interdigitated between penultimate and antepenultimate supralabials; lower eyelid scaly, moveable, with a clear palpebral disc approximately size of ear opening, scales of upper palpebral series 8–11, of lower palpebral series 13–14; supraciliaries eight (n = 2), nine (n = 7), or ten (n = 3), first in contact with prefrontal, last expanded medially behind posterolateral margin of fourth supraocular, none interdigitated with supraoculars; primary temporal single, interdigitated between last two supralabials; secondary temporals two, upper approximately same size as lower but more elongate and overlapping it; supralabials six (n = 1) or seven (n = 11), fourth (n = 1) or fifth (n = 11), respectively, below eye and in contact with small scales of lower eyelid; postlabials two; ear opening moderately large, with 3–6 flat lobules on anterior margin, otherwise ear margin surrounded by flat scales.
Mental single; postmental single, contacting first two infralabials; infralabials seven; four pairs of enlarged chin shields, the first two pairs in medial contact, third pair narrowly separated by single medial scale, fourth pair separated by three medial scales; a single postgenial separating fourth chin shield from seventh infralabial.
Body scales smooth, in 30 (n = 4), 31 (n = 1), or 32 (n = 1) longitudinal rows at midbody; paravertebral scales 52 (n = 3), 53 (n = 1), or 54 (n = 2); medial precloacal scales overlap lateral precloacals.
Scales on dorsal surface of fourth toe in two rows proximally, in single row beginning after last joint, 3 (n = 8) or 4 (n = 4) single dorsal scales; 22–28 smooth subdigital lamellae under fourth toe (mean = 23.2, SD = 0.463, n = 12).
In preservative, dorsal ground color brown (n = 2) or greenish brown (n = 2), with two rows of large, darkbrown mid-dorsal spots, sometimes connected medially, 13–16 spots in each row; additional dorsolateral row of similar spots on each side ( Fig. 2A,B View FIGURE 2 ); sides with several rows of smaller dark-brown spots; many scales in dorsal and lateral ground color laterally margined in dark brown, giving the appearance of narrow dark-brown longitudinal lines on dorsum and sides ( Fig. 2B View FIGURE 2 ). Dorsal surface of tail with single medial row of 6–8 large dark-brown spots proximally, changing to two rows of smaller, less-well-developed dorsolateral spots distally. Limbs and digits heavily spotted with dark brown dorsally. Ground color of head darker brown than on body, heavily marked with dark-brown along most scale margins. Ventral surfaces of body and limbs uniform white, but flecked or dusted with brown on chin, sides of neck, and under tail.
In life, BPBM 18691 and 18692 had light-brown dorsal and lateral scales with dark-brown margins, giving the appearance of stripes; two paravertebral rows of black squares ( Fig. 3A View FIGURE 3 ); sides with 3–4 rows of light dashes; ground color on sides darker than on dorsum; venter white with a green sheen; palms and soles dark yellow; iris dark. The venter of BPBM 18692 was also white (no green sheen noted), but that of BPBM 18693 was yellow.
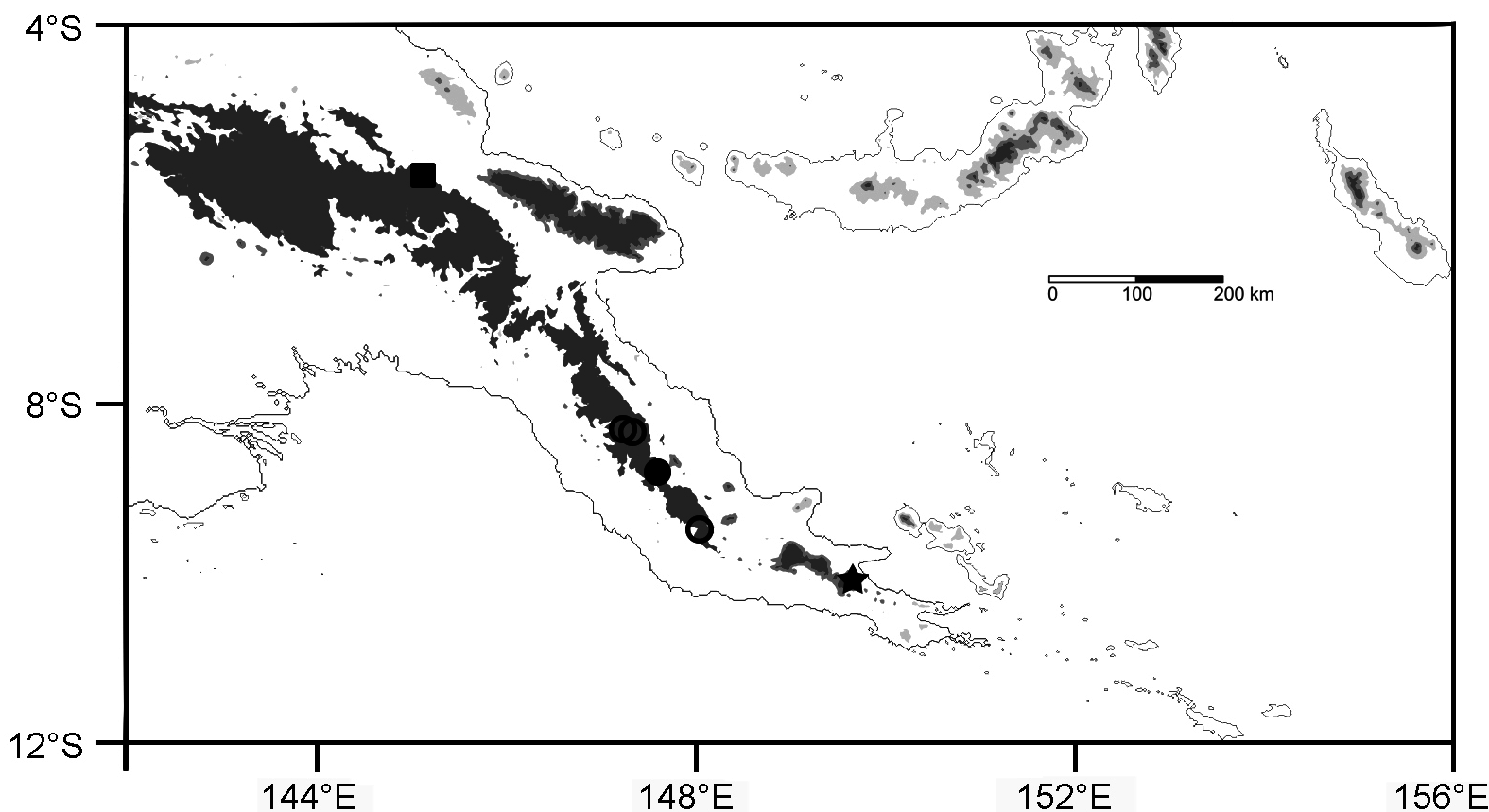
Range. Known with certainty only from four localities in the southern slope of the Owen Stanley Mts. , Central Province, Papua New Guinea ( Fig. 4 View FIGURE 4 ). Elevations for the BMNH specimen from Mt. Victoria is unknown. The specimens from Fane were obtained from 1400–1440 m a.s.l. ; the specimen from Mondo (ca. 5 km E of Fane) was collected at 1520 m a.s.l. , and the specimen from Mt. Obree came from 1330–1450 m a.s.l.
Ecological notes. My specimens were brought in by local collectors, so I have no detailed ecological notes for them. The species is reported to be arboreal ( Greer 1974; Allison 1980), but those statements likely ( Greer 1974) or certainly ( Allison 1980) belong to other, unnamed, though closely related, species (see Discussion). Nonetheless, the flattened body and long digits of L. elegans are consistent with the interpretation that the species is arboreal. The areas from which the animals at Fane and Mt. Obree were collected comprised a mix of open grasslands (likely due to human burning) and secondary forest ( Fig. 5A View FIGURE 5 ), suggesting that this species is heliothermic and prefers areas open to sunshine for basking. It may well be an ecotone specialist.
Remarks. As briefly noted in the Introduction, the large majority of specimens assigned to this species in museum collections I have seen are referable to other, mostly undescribed species that differ from true Lobulia elegans in a variety of scalational and color-pattern features (see Discussion for further details). However, only one of these populations currently has a binomen in synonymy with Lobula elegans :
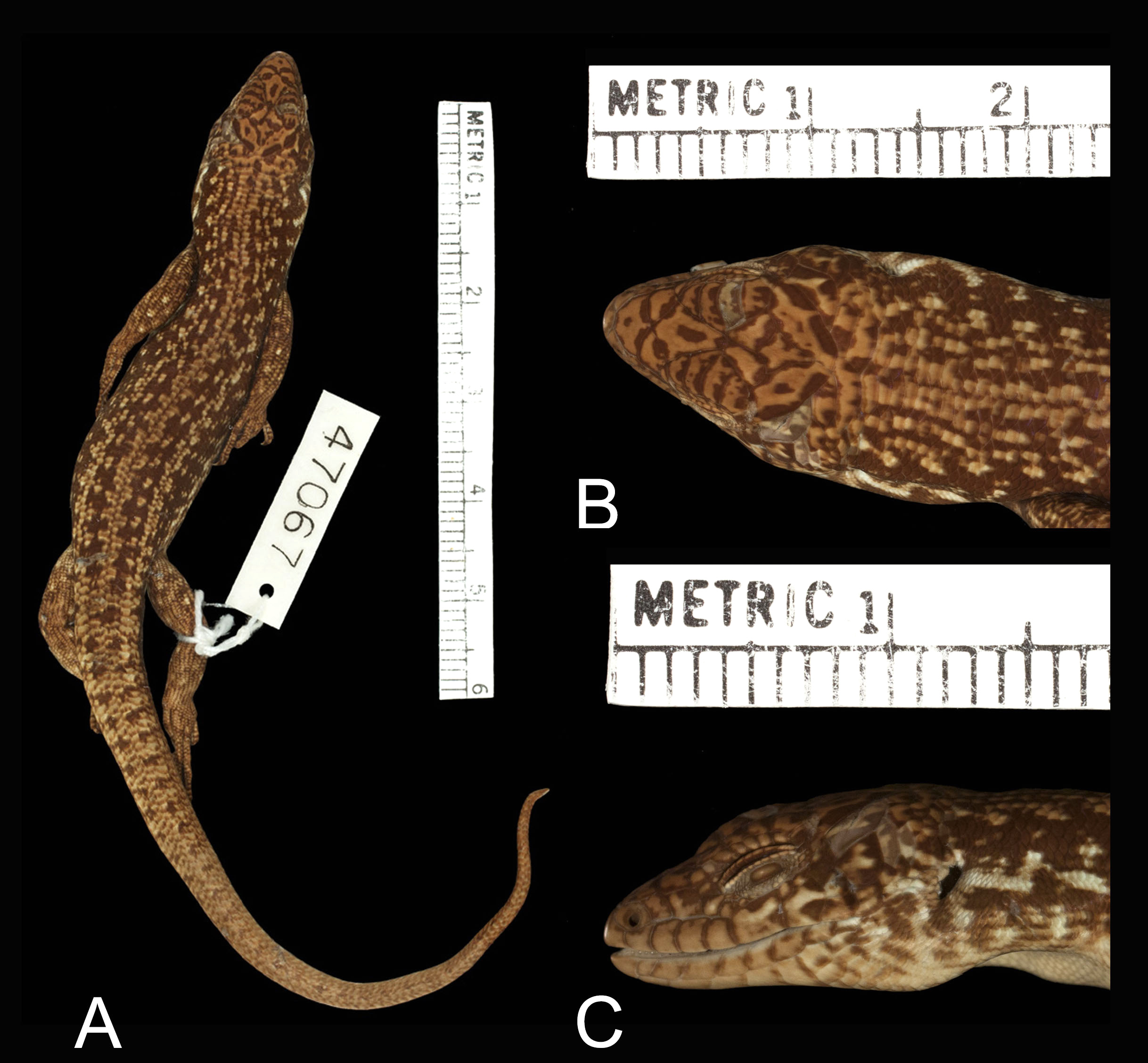
Lygosoma elegantoides lobulus was described by Loveridge (1945) from 2280–2440 m a.s.l., Mt. Wilhelm, Madang Province, PNG, but it seems not to have been mentioned again in the scientific literature, although it is included in the Reptile Database as a subspecies of L. elegans ( Uetz et al. 2019) . The holotype is MCZ 47067 and clearly does not represent the same species as L. elegans as diagnosed here. Based on information provided in Loveridge (1945) and photographs of the holotype available from MCZ ( Fig. 6 View FIGURE 6 ), it differs from L. elegans in having 34–36 midbody scale rows (vs. 30–32 in L. elegans ), posterior loreal square or slightly higher than long (vs. longer than high in L. elegans ), dorsal rows of dark-brown spots joined to form two mid-dorsal stripes (vs. dorsal spots distinct and not arrayed in stripes in L. elegans ), white lateral stripe present (vs. absent in L. elegans ), and paired dark-brown dorsal spots on the tail (vs. single row in L. elegans ). Examining the 16 specimens that comprise the type series of this taxon is beyond the scope of this study, but it is clear from consideration of the above-mentioned features that this taxon is not conspecific with L. elegans , and I hereby remove it from the synonymy of that species and recognize it as the distinct species, L. lobulus .
Ahl (1925) proposed the replacement name Lygosoma elegantoides for Boulenger’s Lygosoma elegans , claiming that the name was preoccupied by both Hinulia elegans Gray and Euprepes (Tiliqua) elegans Fischer once Boulenger (1887) placed 43% of global skink diversity in the single genus Lygosoma . This replacement name has been little used in taxonomic works since then, with L. elegans being used by Greer (1974), Zweifel (1980), Allison & Greer (1986), Mys (1988), and Greer et al. (2005), and L. elegantoides used by Loveridge (1945) and Mittleman (1952). It is well beyond the scope of the present taxonomic work to wade into the wake of nomenclatural problems created by Boulenger’s reconceived Lygosoma , and I leave that to skink experts, who can do a more thorough job. Meanwhile I use the name Lobulia elegans herein simply to be consistent with all recent taxonomic literature for the species.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lobulia elegans (Boulenger)
| Kraus, Fred 2020 |
Lobulia elegans
| Greer, A. E. Jr. 1974: 9 |
Scincella elegantoides
| Mittleman, M. B. 1952: 24 |
Lygosoma (Leiolopisma) elegans, Smith 1937: 224
| Smith, M. A. 1937: 224 |
Lygosoma elegantoides
| Ahl, E. 1925: 20 |
Lygosoma elegans
| Boulenger, G. A. 1897: 8 |