
Mantidactylus ulcerosus (Boettger, 1880)
|
publication ID |
https://doi.org/ 10.11646/megataxa.7.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:2FD8C310-6486-4592-92F6-5EB894EBD6AC |
|
DOI |
https://doi.org/10.5281/zenodo.7504358 |
|
persistent identifier |
https://treatment.plazi.org/id/5F25F715-FF8E-FF92-4F13-4E2F48597C2E |
|
treatment provided by |
Plazi |
|
scientific name |
Mantidactylus ulcerosus |
| status |
|
Mantidactylus ulcerosus View in CoL clade
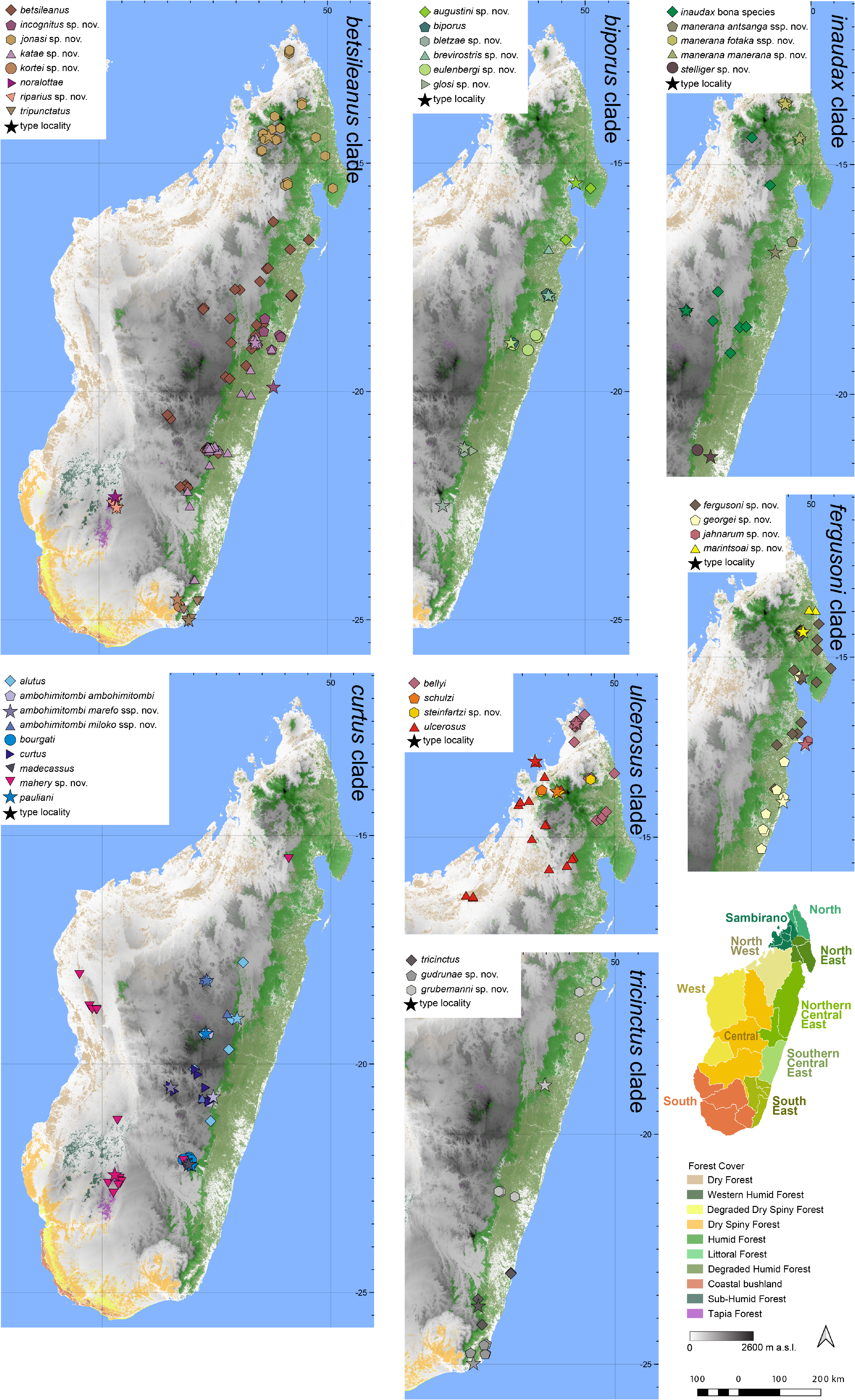
This clade contains two species characterized by a rather stout morphology and medium to large body sizes (28.5– 46.4 mm adult SVL), occurring in the North, Sambirano, and North West regions, and from some sites in the North East ( M. ulcerosus , M. bellyi ) as well as two small-sized species (17.3–28.9 mm adult SVL) apparently restricted to the Tsaratanàna and Manongarivo Massifs in the Sambirano Region ( M. schulzi , and one new species named herein as M. steinfartzi sp. nov., based on the holotype depicted in Fig. 23 View FIGURE 23 ). Mantidactylus ulcerosus ( Boettger, 1880)
Type material.—The taxon Limnodytes ulcerosus Boettger, 1880 is based on a syntype series from ‘ insula Nossi-Bé’ that according to Frost (2021) included SMF 1068.1a–b , MCZ 9331–9334 and 2164 (on exchange from SMF; Barbour & Loveridge 1929), and UMMZ 60296 (on exchange from MCZ). The name-bearing type has been considered to be SMF 6605 , lectotype designated by implication as it was considered the holotype by Mertens (1967), Guibé (1978) and Blommers-Schl̂sser (1979). However, in the original description, Boettger (1880) mentioned explicitely a male and a female specimen, and provided morphometric data for these two specimens. Also, he stated ‘multa spec.’ after the locality information indicating that his description was based on more than one specimen. Mertens (1967) and subsequent authors therefore must have been aware of the fact that Limnodytes ulcerosus was based on syntypes.
Furthermore, Article 74.5 of the Code ( ICZN 1999) states unambiguously for lectotype designations before 2000, that ‘When the original work reveals that the taxon had been based on more than one specimen, a subsequent use of the term “ holotype ” does not constitute a valid lectotype designation unless the author, when wrongly using that term, explicitly indicated that he or she was selecting from the type series that particular specimen to serve as the name-bearing type.’ Since the lectotype designation by the authors listed above was not explicit in terms of the Code, we here stabilize it by expressly designating SMF 6605 (from which we obtained a 16S sequence) as lectotype of Limnodytes ulcerosus out of the available series of syntypes, in agreement with the choice of this specimen by previous authors.
Identity.—Only one lineage of Brygoomantis has so far been reliably recorded from Nosy Be ( Andreone et al. 2003; and the present study), and the nomen ulcerosus has been correctly assigned to this lineage in the recent literature (e.g. Glaw & Vences 1992a, 1994, 2007; Penny et al. 2017; Perl et al. 2014; Vieites et al. 2009).
This assignment was confirmed by the 16S sequence of the lectotype, obtained by barcode fishing and included in our molecular analysis. Genetic results confirmed that this species occurs in the Sambirano region, and at Sahamalaza in the North West ( Penny et al. 2017), across the northern mountain escarpment from Tsaratanàna to Makira. However, all records from eastern Madagascar (e.g. Blommers-Schl̂sser & Blanc 1991) belong to other, unrelated species of Brygoomantis .
Synonyms.—The nomen Mantidactylus brauni Ahl, 1929 is usually considered a junior synonym of Mantidactylus biporus (e.g. Frost 2021; Guibé 1978) but has been considered doubtfully distinct from that species by Blommers-Schl̂sser and Blanc (1991), and as a nomen dubium by Glaw and Vences (1992a). It is based on two unnumbered syntypes from ‘Akkoraka (Central-Madagascar)’ according to the original description, and on a ‘holotype’ ZMB 31617 (a supposed lectotype designation ‘by implication’) by Guibé (1978).
However, Blommers-Schl̂sser & Blanc (1991) doubted whether the syntypes of this species have been correctly identified, and furthermore, as discussed above, a lectotype designation by implication is not a valid nomenclatural act according to article 74.5 of the Code.
We located the specimen ZMB 53737 (locality according to catalogue: ‘Akkorotha, Madagaskar’; collector / donor: [S.G.] Braun) at the Museum f̧r Naturkunde (Berlin) labelled as type of M. brauni and succeeded to obtain a 16S sequence by barcode fishing from this specimen. Consequently, we here deviate from Guibé (1978) and designate specimen ZMB 53737 as lectotype of Mantidactylus brauni Ahl, 1929 , which is justified in order to clarify the identity of this nomen. According to the respective 16S sequence, the lectotype firmly cluster among sequences of M. ulcerosus , and based on this genetic information we include Mantidactylus brauni as junior synonym of M. ulcerosus .
Diagnosis.—A member of the M. ulcerosus clade as revealed by the phylogenomic analysis, and sister to the morphologically very similar M. bellyi . See Table 4 View TABLE 4 for a list of diagnostic morphological characters. The combination of a large body size of up to 45 mm, strongly tubercular dorsal skin usually without welldefined dorsolateral ridges, large tympanum size in males (11–14% of SVL), absence of white spots on flanks and of white marking on snout tip, and pulsed advertisement calls emitted in regular series distinguishes M. ulcerosus from species of the other clades. Some species in the M. fergusoni clade can be morphologically similar, but they occur in eastern Madagascar (vs Sambirano and North West regions), and have highly different advertisement calls ( Table 4 View TABLE 4 ). Within the M. ulcerosus clade, the new species differs by its large body size and tubercular dorsal skin from M. schulzi , and by its advertisement call emitted in regular call series from M. bellyi (single calls). M. ulcerosus can occur sympatrically with M. mahery in the North West of Madagascar, but that species differs by a smoother skin and fewer pulses per note in advertisement calls. For detailed distinction from new species described herein, see the respective species accounts. A full list of molecular diagnostic sites in the 16S gene of M. ulcerosus in pairwise comparisons to all other Brygoomantis species is provided as Supplementary appendix.
Re-description of the lectotype (SMF 6605). Adult female in good state of preservation. Body stout. Head as wide as body. Snout rounded. Nostrils directed laterally, slightly protuberant. Nostrils nearer to tip of the snout than to eye. Canthus rostralis weak, slightly concave. Loreal region weakly concave. Tympanum distinct, large, rounded, diameter about 68% of eye diameter. Supratympanic fold distinct, beginning straight, with a rather distinct bend midway towards insertion of forelimb. Tongue ovoid, distinctly posteriorly bifid. Maxillary teeth present.Vomerine teeth present in two rounded aggregations, positioned posterolateral to choanae. Choanae rounded.
Fingers without webbing. Relative length of fingers: I<II<IV<III. Finger discs slightly enlarged. Nuptial pads absent. Subarticular tubercles single. Outer metacarpal tubercle not recognisable, inner metacarpal tubercle present. Foot almost as long as tibia (96%). Lateral metatarsalia separated. Inner metatarsal tubercle present. Outer metatarsal tubercle present. Webbing formula: 1(0.5), 2i(1), 2e(0.5), 3i(2), 3e(1), 4i(2), 4e(1.5), 5(0). Relative length of toes: I<II<V<III<IV. Skin on the upper surface with few scattered granules and tubercles on flanks. Ventral side smooth. Femoral glands small but present. Proximal granular gland field present.
Colour in preservative: dorsum red-brown, with indistinct irregular darker markings. Forelimbs light brown with poorly defined darker markings. Hindlimbs light brown with indistinct darker crossbands. Inguinal region without few scattered whitish spots. Snout tip without a whitish spot. Venter beige, throat darker than belly. Lower lip with distinct irregular brown spots. Toe discs light brown to grey. Inner side of tibia brown mottled with beige.
Variation.—Variation in measurements is given in Table 6 View TABLE 6 . See Fig. 24 View FIGURE 24 for colouration in life and its variation.
There is moderate sexual size dimorphism (confirmed male SVL 28.5–35.9 mm [n = 8] vs confirmed female SVL 32.5–45.4 [n = 14]). Males have a larger tympanum than females (HTD/ED ratio is 53–79% in females, 74– 95% in males). Specimens with a light vertebral stripe, or with a broad light vertebral band, occur regularly.
Dorsolateral ridges are absent in many individuals, but can be recognised in others ( Fig. 24 View FIGURE 24 ). Femoral glands in males are distinct, often orange-coloured in life, and with distinct distal ulcerous macrogland and proximal granular gland field; the granular gland fields on the two opposite thighs contact each other medially.
Natural history.—Common along small streams with shallow water and associated swamps, where males emit their advertisement calls at night from the ground next to the water or sitting in shallow water. Males often begin calling in the late afternoon. Found in primary rainforest or transitional/dry forest and also in degraded areas and secondary forest, as long as some vegetation cover is present around the streams. Previous descriptions of eggs and embryos (Blommers-Schl̂sser 1979) refer to other species of Brygoomantis . Figure 24n View FIGURE 24 shows a clutch of eggs probably assignable to this species.
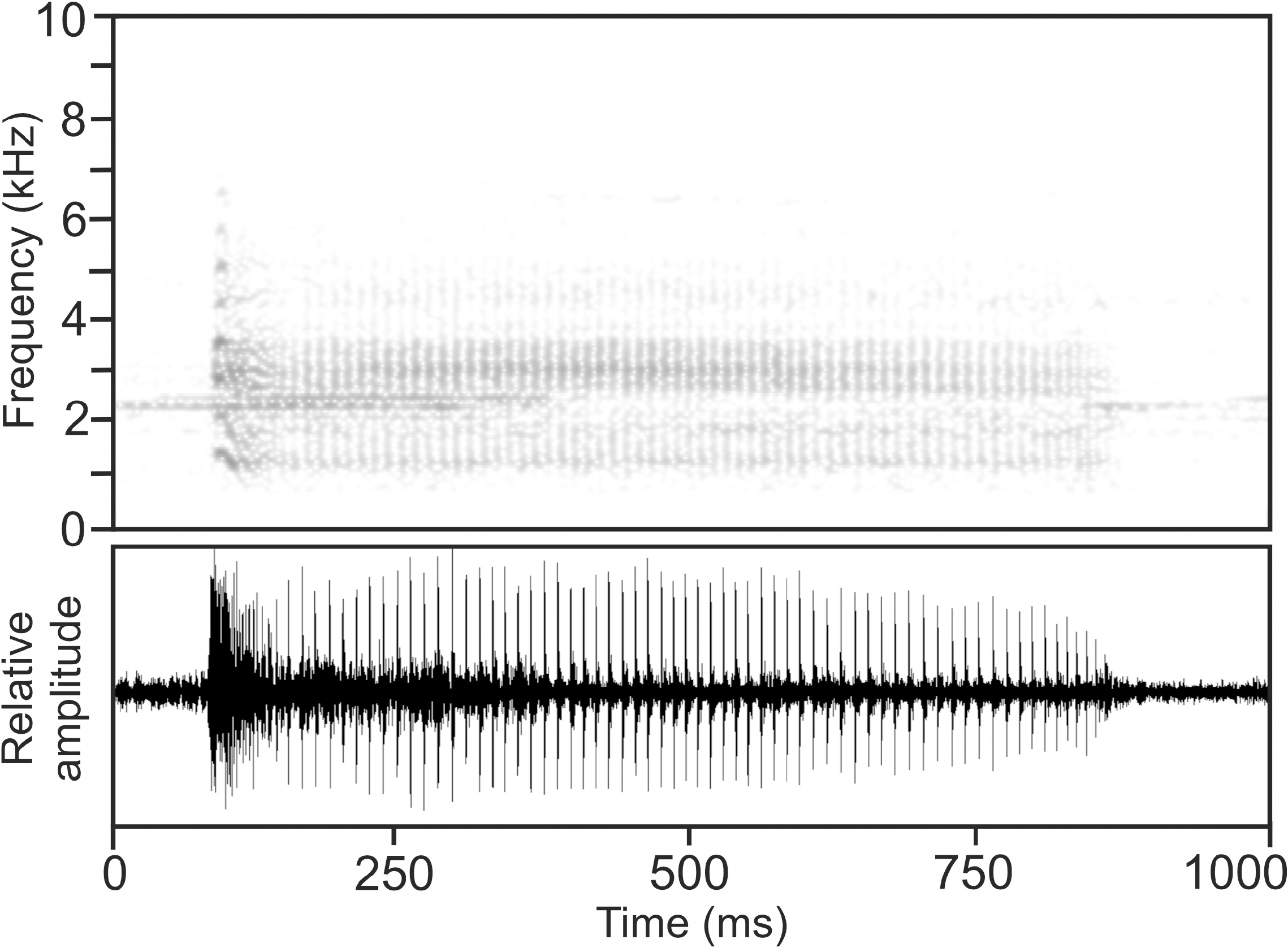
Calls.— The advertisement call of M. ulcerosus , recorded on 10 February 1992, 19:00 h, at Nosy Be ( Vences et al. 2006: CD 2, track 74, cut 1), consists of a pulsed note ( Fig. 25 View FIGURE 25 ), emitted in series at regular intervals and very fast succession (short inter-call intervals).
Notes exhibit some complexity in pulse structure, with the initial pulse of each note being distinctly longer in duration and containing highest call energy compared to subsequent pulses. This initial pulse seems to contain some substructure and amplitude modulation and sounds more distorted than subsequent pulses. Relative amplitude of pulses slightly decreases towards the end of the note.
Within the recorded short call series of four calls, these become successively shorter from the beginning to the end of the series. Numerical parameters of four analysed calls were as follows: call duration (= note duration) 453– 779 ms (593.0 ± 143.2 ms); 45–65 pulses per note (54.8 ± 9.0); pulse duration 1–3 ms (2.3 ± 0.6 ms); pulse duration of initial pulses 27–37 ms (31.8 ± 4.1 ms); pulse repetition rate within notes (excluding initial pulse) 83.9–104.5 pulses/s (92.4 ± 7.6); dominant frequency 3101–3605 Hz (3308 ± 196 Hz); prevalent bandwidth 880–5500 Hz; call repetition rate (= note repetition rate) within regular call series ca 68 calls/min.
Calls recorded from a chorus on 30 June 2009 from a site at km 27 of the road from Antsohihy to Mandritsara, 24°C air temperature, generally agreed in character with calls described from Nosy Be. They also exhibit the complex pulse structure described above, with initial pulses being very narrowly spaced and sometimes fused.
Calls were emitted in series, containing 4–9 calls. The recording was difficult to analyse due to many overlapping calls, but the following call parameters could be measured (10 calls analysed): call duration (= note duration) 355– 565 ms (447.4 ± 63.0 ms); pulse duration 1–4 ms (1.8 ± 0.9 ms); pulse repetition rate within notes varied from ca 50–220 pulses/s; dominant frequency 2798–2885 Hz (2847 ± 37 Hz); prevalent bandwidth 1000–3800 Hz; call repetition rate (= note repetition rate) within regular call series ca 70–88 calls/min.
Calls recorded on 24 February 2001 atAnkarafantsika, 28°C air temperature, also agreed with the calls described above, with very narrowly spaced initial pulses, partly fused. Calls were emitted in series containing 3–5 calls.
Recordings were of poor quality, containing overlapping calls and background noise. However, the following parameters could be measured (8 calls analysed): call duration (= note duration) 551–1047 ms (667.3 ± 160.0 ms); pulse repetition rate within notes varied from ca 50–400 pulses/s (maximum values from initial pulses); prevalent bandwidth 900–3400 Hz; call repetition rate (= note repetition rate) within regular call series ca 66–87 calls/min.
Distress calls of a female from Nosy Be were described by Glaw and Vences (1992b).
Tadpoles.— Tadpoles assignable to this species (from the type locality Nosy Be) were briefly mentioned and their tooth formula described by Glaw & Vences (1994).
The tadpole description by (Blommers-Schl̂sser 1979) was based on east coast specimens not identified by genetics that almost certainly belong to other species of Brygoomantis .
Distribution.— Widespread in the North West (including the western slopes of the Makira Reserve) and Sambirano regions of Madagascar, over a wide range of elevations and habitat types ( Fig. 7 View FIGURE 7 ). This species is known from Angorony, Ankarafantsika, Antsatramidola, Benavony, Berara, the border of the Bealanana district (Bevitagnono and Irogno forest), between Antsohihy and Mandritsara, between Antoshihy and Port Berger, Makira West (Sahaovy, Camp 0), Nosy Be (type locality) including Lokobe National Park, Sahamalaza, and Tsaratanàna. It was also recorded from Nosy Komba ( Hyde Roberts & Daly 2014). Records from the forests of Belambo and Anjiamangirana and in the Namoroka National Park ( Raselimanana 2008) require genetic confirmation. Elevation range: 0–1093 m a.s.l.
Etymology.— Latin adjective meaning ‘full of sores’ or ‘ulcerated’, presumably in reference to the granular dorsal skin or maybe to the femoral glands.


| SMF |
Germany, Frankfurt-am-Main, Forschungsinstitut und Naturmuseum Senckenberg |
| MCZ |
USA, Massachusetts, Cambridge, Harvard University, Museum of Comparative Zoology |
| UMMZ |
USA, Michigan, Ann Arbor, University of Michigan, Museum of Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.