
Nyctibatrachus kumbara, Gururaja, Kotambylu Vasudeva, Dinesh, K. P., Priti, H. & Ravikanth, G., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3796.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:8BA7FB48-FF23-4E83-89B7-BAA040AF215D |
|
DOI |
https://doi.org/10.5281/zenodo.6131139 |
|
persistent identifier |
https://treatment.plazi.org/id/03EB8786-B711-FFF6-D0BB-2E1BFCF7A6CA |
|
treatment provided by |
Plazi |
|
scientific name |
Nyctibatrachus kumbara |
| status |
sp. nov. |
Nyctibatrachus kumbara View in CoL sp. nov.
( Table 2S View TABLE 2 S , Figures 5 View FIGURE 5. a , 6 View FIGURE 6. a , 8–9 View FIGURE 8 View FIGURE 9 , 10 View FIGURE 10 c.)
Holotype. ZSI/ WGFRS /V/A/860, adult male collected on 09-06-2006 by KVG and SA along a flowing rivulet of Sharavathi river basin (14.27323°N; 74.74756°E, 590m amsl) at Kathalekan, Uttara Kannada District, Karnataka, India.
Paratypes. ZSI/ WGFRS /V/A/861 and ZSI/ WGFRS /V/A/863, adult females collected on 03-10-2006 by KVG and SA along a flowing rivulet of Sharavathi river basin (14.27499°N; 74.73695°E, 583m amsl) at Malemane, Uttara Kannada District, Karnataka, India; ZSI/ WGFRS /V/A/862 and ZSI/ WGFRS /V/A/864, adult males collected on 09-06-2011 by KVG and KPD along a flowing rivulet of Sharavathi river basin (14.27323°N; 74.74756°E, 590m amsl) at Kathalekan, Uttara Kannada District, Karnataka, India.
Diagnosis. This species is assignable to the genus Nyctibatrachus because of its semi aquatic to aquatic habitat, medium to large size, pupil rhomboidal, glandular wrinkled skin, presence of vomerine teeth, notched tongue, finger and toe discs with disc, absence of webbing on fingers and presence of webbing on toes and presence of sub ocular gland ( Biju et al. 2011).
Nyctibatrachus kumbara sp. nov. can be easily distinguished from all other species in the genus by the following combination of characters: (1) adult male size medium to large (SVL 46.5± 0.74 mm); (2) head wider than long (HW 18.7–20.9 mm; HL 14.8–15.7 mm); (3) dorsum glandular without any spiny projections in the anterior half, glandular corrugations irregular without specific pattern; in males, throat and chest finely dotted with glandular folds rest of the region smooth, belly white; (4) webbing on toes medium (I ½- 1 II ½- 1 III 0- 2 IV 2-0 V); (5) nuptial pad and femoral glands prominent in adult males; (6) dorsal body color dark brown, ventrally buff colored except belly; (7) finger disc weakly developed (fd3 0.9± 0.24 mm; fw3 0.8± 0.19 mm); (8) toe disc moderately developed (td4 1.7± 0.32 mm; tw4 0.8± 0.05 mm); (9) third finger disc with dorso-terminal groove, cover notched distally, fourth toe disc with dorso-terminal groove cover bifurcate distally; (10) hand-stand oviposition, fertilized egg clutch covered with mud pack; (11) genetically belongs to N. sanctipalustris clade including N. dattatreyaensis (3.41% divergent on mitochondrial 16S rRNA), N. vrijeuni (4.02%), N. shiradi (4.02%), N. karnatakaensis (4.62%) and N. sanctipalustris (4.62%).
Description of the holotype. ZSI/WGRC/V/A/860, adult male ( Figure 9 View FIGURE 9 ), terminology follows Biju et al. (2011) and Dinesh et al. (2008a). Morphometric data are given in Table 2S View TABLE 2 S .
A medium-sized species of Nyctibatrachus (SVL 46.2 mm); habitus compact and squat; head wider (HW 18.7 mm) than long (HL 15 mm; MN 11.9 mm; MFE 9.6 mm; MBE 3.4 mm), rounded in anterior view; snout rounded (SL 7.1 mm), marginally protruding, its length longer than horizontal diameter of eye (EL 5.7 mm); canthus rostralis rounded; loreal region concave; interorbital space flat (IUE 6.3 mm) and double the upper eye lid width (UEW 3.1 mm); internarial distance (IN 4.3 mm) less than interorbital distance (IUE 6.3 mm); distance between back of eyes (IBE 14.5 mm) is greater than double the distance between front of eyes (IFE 6.9 mm); nostrils oval, without a flap of skin, and is closer to eye (EN 2.9 mm) than snout tip (NS 3.5 mm); eyes large (EL/HL = 0.38%) protruding on the sides of head, its diameter more than eye to nostril distance (EL/EN=1.96); pupil rhomboidal; tympanum indistinct; subocular gland distinct ( Figure 9 View FIGURE 9 C); pineal ocellus absent; vomerine ridge present posterior to choanae, oval bearing three on left and six sharp spinose teeth on right ridge; symphysial knob ‘W’ shaped, moderately developed; tongue moderate, oval slightly emarginated, median lingual process absent; parotoid glands, cephalic ridges and co-ossified skin absent.
Forelimbs (FLL 10.9 mm) strong and sub-equal to hand length (HAL 10.1 mm); third finger thin, long, rounded (TFL 5.3mm); thin dermal fringe on finger III; webbing absent ( Figure 9 View FIGURE 9 D); relative length of fingers, shortest to longest: I<II<IV<III; tips of fingers enlarged with weakly developed disc and third finger disc with dorso-terminal groove, cover notched distally ( Figure 9 View FIGURE 9 F); subarticular tubercles moderate, round one each on finger I and II and two each on finger III and IV; prepollex and palmar tubercle distinct ( Figure 9 View FIGURE 9 D).
Hind limbs moderately long; shank length (TL 21.3 mm) more than two times longer than wide (TW 8.1 mm), shorter than thigh length (FL 22.6 mm) and sub-equal to the distance from the base of the internal metatarsal tubercle to the tip of toe IV (FOL 20.1 mm); heels touch when the tibia are folded at right angles to the body; toes thin and long; toe IV long (T4L 9.9 mm) and less than the distance from the base of the tarsus to the tip of toe IV (TFOL 26.7 mm); relative length of toes, shortest to longest: I<II<V<III<IV ( Figure 9 View FIGURE 9 E); tips of toes with moderately developed discs and fourth toe disc with dorsoterminal groove, cover bifurcate distally ( Figure 9 View FIGURE 9 G), webbing medium (3/4th) (I ½- 1 II ½- 1 III 0- 2 IV 2-0 V) (MTTF 13.5 mm; MTFF 12.8 mm; TFTF 6.6 mm; FFTF 8.4 mm) ( Figure 9 View FIGURE 9 H); dermal fringe along toe V distinct; subarticular tubercles present, moderate, oval, one each on toe I and II, two each on toe III and V, and three on toe IV; inner metatarsal tubercle short prominent, spade shaped; its length (IMT 3 mm) shorter than toe I (T1L 3.5 mm) length; outer metatarsal tubercle, supernumerary tubercles and tarsal tubercles absent ( Figure 9 View FIGURE 9 E).
Skin is corrugated on the dorsal and lateral surfaces of the body with glandular folds; forelimbs and hind limbs with short longitudinal glandular folds; snout with a ‘Y’ shaped fold from middle of upper lip, its arms bifurcating between nares and running towards orbits; three small dotted horizontal folds between posterior corner of eyes to region above shoulder; supratympanic fold and subocular fold distinct. Throat with dense glandular longitudinal folds and rest of the venter soft glandular without any folds; belly smooth; lower lip dotted with a single row of small discontinuous glandular folds; thigh ventrally smooth ( Figure 9 View FIGURE 9 B); dorsally tarsus with fine spinules ( Figure 9 View FIGURE 9 A). Distinct raised femoral gland present (FGL 9.2 mm; FGB 3.8 mm).
Color of holotype. In life , iris golden yellow in its upper part, pupil rhomboidal, black in color ( Figure 8 View FIGURE 8 ). Sides of head light colored, region between subocular gland and tympanic region cream colored. A black horizontal band connecting upper eye lids. Dorsum brown with two light brick red dorsolateral bands laterally bordered by dark brown from orbits to vent. Region between dorsolateral bands light brown with three black horizontal bands, one at between orbits, one at anterior region of shoulders and other at posterior region of shoulders. Forearm barred with smalls black and larger dark brown bands; hind limbs barred with small light brick red and larger dark brown bands. Sides of the body brownish. Nuptial pads on base of first fingers cream colored. Throat translucent with fine black spots on the glandular skin. Chest, belly and anterior part of thighs translucent, where internal organs can be clearly seen. Posterior part of thigh with raised glossy light yellow femoral gland in males.
In preservative ( Figure 9 View FIGURE 9 A), dorsal and lateral parts of body mottled with brown and cream color, with a pair of faint dorsolateral lines from posterior orbit to above vent; faint horizontal band between eyes; femoral glands cream white; dorsal parts of limbs, forelimbs, thigh, shank and foot up to tip of fingers and toes barred; throat and chest pale brown; belly and thighs white; femoral glands granular, white in color ( Figure 9 View FIGURE 9 B).
Secondary sexual characters. Males have a nuptial pad covering the base of the dorsal surface of the first finger and a pair of moderately raised femoral glands which are more than two times longer (FGL 9.2 mm) than wide (FGB 3.8 mm), and is conspicuous both in life and preservative ( Figure 9 View FIGURE 9 B). Females have pigmented eggs.
Additional information from paratypes and variations. Morphological data are given in Table 2S View TABLE 2 S . Paratypes range from 42.8 mm to 47.4 mm in SVL; in all the external morphological characters they are similar to holotype.
Etymology. The specific epithet ‘Kumbara’ is derived from Kannada language, a name given to the community of people involved in making pottery. The species name Kumbara is a noun in apposition to generic name.
Suggested common name. Kumbara night frog.
Distribution and Natural History. Distribution of Nyctibatrachus kumbara sp. nov. is here confirmed from the streams and rivulets of the Tunga, Sharavathi, Aghanashini and Bedthi rivers (between 13.60515°N – 14.61582°N and 74.75576°E – 75.320638°E) in Shimoga and Uttara Kannada districts of South India ( Figure 1 View FIGURE 1 ). Based on our field observations, this species is locally abundant in the surroundings of the type locality and is very much restricted to perennial streams of the evergreen and semi-evergreen forests.
Comparison. We compared Nyctibatrachus kumbara sp. nov. with all 27 valid species of Nyctibatrachus based on either the examination of holotypes or non-type vouchers collected from their respective type localities (listed in Appendix 2).
Possession of a combination of characters like; medium to large size; irregular dorsal glandular skin corrugations without specific pattern and spiny projections on the anterior half; third finger disc with dorsoterminal groove, cover notched distally; fourth toe disc with dorso-terminal groove cover bifurcate distally; webbing medium; nuptial pad and femoral glands present in adult males, and egg laying with handstand oviposition and covering of eggs with mud, are unique to Nyctibatrachus kumbara sp. nov.
Detailed morphological comparisons for Nyctibatrachus kumbara sp. nov. is made only for sympatric species. But other allopatric congeners are compared with a suite of characters, namely, SVL, third finger disc, fourth toe disc, femoral gland in male and webbing in foot ( Table 3).
Genetically Nyctibatrachus kumbara sp. nov. belongs to N. sanctipalustris clade ( Figure 7 View FIGURE 7 ). The other species included in the N. sanctipalustris clade differ from the Nyctibatrachus kumbara sp. nov. as follows: N. dattatreyaensis is smaller (SVL 36.2 to 42.3 mm), third finger disc without dorso-terminal grove, fourth toe disc with dorso-terminal grove cover notched distally; N. vrijeuni is smaller (SVL 37.4 to 43.1 mm), third finger disc without dorso-terminal grove, fourth toe disc with dorso-terminal grove cover notched distally; N. shiradi is smaller (SVL 18.8 to 27.5 mm), with small webbing, fourth toe disc with dorso-terminal grove cover notched distally; N. sanctipalustris Is smª11er (SVL 25.7 to 37.6 mm), third finger disc without dorso-terminal grove, fourth toe disc with dorso-terminal grove cover notched distally; N. karnatakaensis is larger (SVL 56.0 to 63.8 mm), with extensive webbing, third finger disc with dorso-terminal grove cover rounded distally, fourth toe disc with dorsoterminal grove cover rounded distally. Nyctibatrachus kumbara sp. nov. exhibited 3.41–4.62% genetic divergence to these species ( Table 1 View TABLE 1 S).
Nyctibatrachus kumbara sp. nov. is syntopic with N. kempholeyensis ( Figure 10 View FIGURE 10 a) and N. jog ( Figure 10 View FIGURE 10 b) but can be distinguished in the field from N. kempholeyensis in having medium to large adult size SVL 42.8 to 47.4 mm (vs small adult size SVL 15.5 to 24.4mm in N. kempholeyensis ), finger and toe disc weak to moderately developed (vs finger and toe disc well developed in N. kempholeyensis ), glandular dorsum without any spiny projections in the anterior half, glandular corrugations irregular without specific pattern (vs less wrinkled dorsal skin with prominent granular projections in N. kempholeyensis ), webbing medium (vs webbing small in N. kempholeyensis ), egg clutches covered with mud (vs eggs without mud packing in N. kempholeyensis ), eggs are laid at the edges of the slow flowing streams (vs eggs are laid at the slushy edges of steams in N. kempholeyensis ).
Nyctibatrachus kumbara sp. nov. can be distinguished in the field from N. jog in having medium to large adult size SVL 42.8 to 47.4 mm (vs medium adult size SVL 33.1 to 39.0 mm in N. jog ), finger and toe disc weak to moderately developed (vs finger and toe discs well developed in N. jog ), glandular dorsum without any spiny projections in the anterior half, glandular corrugations irregular without specific pattern (vs relatively weakly wrinkled dorsal skin with prominent glandular projections in N. jog ), egg clutches with 8 eggs and are covered with mud (vs egg clutches with 52 eggs without mud packing in N. jog ), eggs are laid at the edges of the slow flowing streams (vs eggs are laid at the edges and above the flowing streams in N. jog ).
Reproductive character displacement. Reproductive characteristics of 13 species of Nyctibatrachus are presented in Table 4. Detailed reproductive characteristics were available for N. humayuni , N. petraeus , N. jog ( N. humayuni clade, spatially non-overlapping and allopatric, as evidenced from the genetic tree in Figure 7 View FIGURE 7 ), N. kempholeyensis and Nyctibatrachus kumbara (spatially overlapping and sympatric). On comparing reproductive characters between these two groups, we found less difference among the allopatric species and significant difference between the sympatric species with respect to SVL ( Figure 11 View FIGURE 11 ). Table 5 View TABLE 5 details unpaired one tailed T test to compare allopatric and sympatric species. The remaining reproductive characters were subjected to non-metric magnification under 1.0x objective)).
Species SVL(mm, *Third finger disc *Fourth toe disc Femoral Webbing
maximum size of gland in adult ♂and♀) male
N. acanthodermis 49.7 to 66.2 without groove with dorso-terminal groove, cover Present Medium
rounded distally
N. aliciae 20.5 to 32.0 with dorso-terminal groove, with dorso-terminal groove cover Present Medium
cover notched distally bifurcate distally
N. anamallaiensis 13.1 to 17.4 with dorso-terminal groove with dorso-terminal groove cover Present Absent
cover bifurcate distally bifurcate distally
N. beddomii 13.3 to 18.0 with dorso-terminal groove with dorso-terminal groove cover Present Absent
cover bifurcate distally bifurcate distally
N. danieli 24.9 to 35.1 with dorso-terminal groove with dorso-terminal groove cover Absent Medium
cover rounded distally rounded distally
N. dattatreyaensis 36.2 to 42.3 without dorso-terminal groove with dorso-terminal groove, cover Present Medium
notched distally
N. deccanensis 17.4 to 26.2 without dorso-terminal groove with dorso-terminal groove, cover Present Rudiment
bifurcate distally
N. deveni 22.6 to 33.6 with dorso-terminal groove, with dorso-terminal groove, cover Absent Medium
cover notched distally bifurcate distally
N. gavi 49.5 to 60.1 without dorso-terminal groove with dorso-terminal groove, cover Present Medium
rounded distally
N. grandis 56.8 to 76.9 without dorso-terminal groove with dorso-terminal groove, cover Present Medium
rounded distally
N. humayuni 32.8 to 50.6 with dorso-terminal groove with dorso-terminal groove cover Present Medium
cover rounded distally rounded distally
N. indraneili 42.5 to 50.8 without groove without groove Present Medium
N. jog 33.1 to 39.0 with dorso-terminal groove with dorso-terminal groove cover Present Medium
cover rounded distally rounded distally
N. karnatakaensis 56.0 to 63.8 with dorso-terminal groove with dorso-terminal groove cover Present Extensive
cover rounded distally rounded distally
N. kempholeyensis 15.5 to 24.4 with dorso-terminal groove, with dorso-terminal groove, cover Present Small cover rounded distally rounded distally
……continued on the next page Species SVL(mm, *Third finger disc *Fourth toe disc Femoral Webbing
maximum size of gland in adult ♂and♀) male
Nyctibatrachus kumbara sp. 42.8 to 47.4 with dorso-terminal groove with dorso-terminal groove, cover Present Medium
nov. cover notched distally bifurcate distally
. major 31.5 to 54.2 without dorso-terminal groove with dorso-terminal groove, covers Present Medium
rounded distally
. minimus 10.0 to 14.8 with dorso-terminal groove, with dorso-terminal groove, cover Present Absent
cover bifurcate distally bifurcate distally
. minor 15.4 to 20.5 with dorso-terminal groove with dorso-terminal groove cover Absent Absent
cover bifurcate distally bifurcate distally
. periyar 24.2 to 29.9 with dorso-terminal groove, with dorsoterminal groove, cover Present Medium
cover notched distally bifurcate distally
. petraeus 35.1 to 47.6 with dorso-terminal groove with dorso-terminal groove cover Present Medium
cover rounded distally rounded distally
. pillaii 20.0 to 25.4 with dorso-terminal groove with dorso-terminal groove cover Absent Medium
cover bifurcate distally bifurcate distally
. poocha 25.3 to 37.5 with dorso-terminal groove with dorso-terminal groove cover Present Medium
cover bifurcate distally bifurcate distally
. sanctipalustris 25.7 to 37.6 without dorso-terminal groove with dorso-terminal groove, cover Present Medium
notched distally
. shiradi 18.8 to 27.5 with dorso-terminal groove with dorso-terminal groove cover Present Small
cover notched distally notched distally
. sylvaticus 27.5 to 36.2 with dorso-terminal groove with dorso-terminal groove cover Present Medium
cover rounded distally rounded distally
. vasanthi 21.9 to 37.0 with dorso-terminal groove with dorso-terminal groove cover Present Medium
cover bifurcate distally bifurcate distally
. vrijeuni 37.4 to 43.1 without dorso-terminal groove with dorso-terminal groove, cover Present Medium
notched distally
multidimensional analysis. Figure 12 View FIGURE 12 illustrates the NMDS plot based on SVL (male and female), maximum number of eggs in a clutch, type of amplexus (absence=0, loose abbreviated=1, axillary=2), parental care (absent=0, by male=1, by both male and female=2), mud covering behavior (absence=0, presence=1), oviposition site (<10cm from water=1,>10 and <30 cm from water=2,> 30cm from water=3) and position of male and female at the time of oviposition (upright=1, handstand=2) among the overlapping and non-overlapping groups (stress value = 0.11). There is a clear clustering of allopatric species of Nyctibatrachus to sympatric species ( Figure 12 View FIGURE 12 ).
For the co-occurring congeneric (COCG) species call characteristic features were subjected to statistical analysis. Overall call characteristic features (duration and peak frequency) between all COCG species were significantly different (ANCOVA, F=669.1, P<0.00001). On pairwise comparison of call characterstics of COCG species, except for N. jog and N. kumbara (ANCOVA, F<1, p>0.5), remaining COCG species exhibited significant difference ( Table 6 View TABLE 6 ). Amplexus in N. jog is loose amplexus, while in N. kempholeyensis (KVG personal observation) and in N. kumbara it is axillary. Oviposition site for N. jog is wet rock surfaces, overhanging leaves and tree bark above 15 cm to ~ 200 cm from the bottom of stream, but it was only 4−10 cm for N. kumbara and within 30 cm for N. kempholeyensis . Egg clutch size significantly differed among COCG species ( Table 5 View TABLE 5 ).
Most significant reproductive character displacement is in terms of parental care. Nyctibatrachus kumbara covers eggs with mud, while N. jog and N. kempholeyensis do not cover the eggs, but attend the egg clutches. In terms of breeding period, again there is an overlap between N. kumbara and N. kempholeyensis , but they differ from N. jog which breeds only during monsoon (KVG personal observation).
Anurans exhibit diverse reproductive strategies to overcome the limitation posed by external or internal fertilisation ( Beck 1998), selective pressure of predators on aquatic eggs and tadpoles (Prado et al. 2005), prevailing environmental conditions both in space and time; and as a part of evolutionary process ( Jameson 1955; Duellman & Trueb 1994; Wells 2007; Pfenning & Stewart 2010). Also r- and K-selection (MacArthur & Wilson 1967) influence the reproductive strategies in anurans ( Nichols et al. 1976). However, according to Stearns (1992) the concept of r- and K-selection is less meaningful due to varied range in life-history strategies seen in nature and does not make precise predictions. In the present study, we compared three co-occurring species of Nyctibatrachus , namely N. jog , N. kempholeyensis and N. kumbara and three allopatric species, namely N. humayuni , N. petraeus and N. jog ( Figure 10 View FIGURE 10 ). Nyctibatrachus jog is alloptric with the N. humayuni clade, however it is sympatric with N. kempholeyensis and N. kumbara .
Significance of courtship. Mate identification through tactile cues is an important non-vocal signal in anurans ( Duellman & Trueb 1994). Male individuals of N. kumbara called from a fixed site and amplected with an approaching female, which is similar to the species of Bombina , Discoglossus , Pelobates , Scaphiopus , Bufo and Rana ( Wells 2007) . In N. kumbara the courtship behaviour exhibited by males and females by touching each other is perhaps to recognise the potential mate and to gauge the size of both individuals. However, all this remain very speculative and calls for further studies to ascertain the behaviour.
Significance of mud pack on egg clutches. Parental care in amphibians is the energy investment into offspring after fertilisation (for details refer to Wells 2007). Egg attendance by male individuals has been attributed to protection of eggs from being damaged by intruding males, and deterring egg predation and preventing desiccation (in Hyalinobatrachium valerioi ( Vockenhuber et al. 2009)) . Parental care involving both male and female, only females and only males are well known ( Wells 2007). So far, there is no report of any other anuran species covering egg clutch with mud. Mud packing on egg clutches by N. kumbara might be attributed to, 1. minimising clutch dehydration as the outer capsule of terrestrial amphibian eggs are susceptible to dehydration by losing moisture to drier air ( Duellman & Trueb 1994), 2. camouflage eggs as the eggs are very prominently pigmented to avoid predators ( Crump 1995; Wells 2007).
Reproductive Character Displacement. Observed reproductive characters in N. jog , N. humayuni and N. petraeus are similar with loose to abbreviated amplexus ( Kunte 2004; Narahari et al. 2011; Biju et al. 2011, detailed in Table 4). They belong to the N. humayuni clade from the Northern Western Ghats ( Bocxlaer et al. 2012 and Figure 7 View FIGURE 7 ). The minimal aerial distance between the known populations of this clade are N. jog-N. petraeus is 97 km; N. jog-N. humayuni is 331 km and N. petraeus-N. humayuni is 164 km (distance calculated from the nearest verified population of the respective species using Google Earth). However, information on reproductive characters from other clades of Nyctibatrachus from the Western Ghats is lacking. Despite such limitation, on comparing reproductive characters between spatially non-overlapping allopatric to overlapping sympatric clades, we found differences in reproductive characters among the spatially overlapping taxa and reproductive character similarities in spatially non-overlapping taxa in a suite of character, which could be an indication for reproductive character displacement.
Significance of new species. Taxonomy for the genus Nyctibatrachus has been stabilised in the recent past ( Das & Kunte 2005; Dinesh et al. 2007; Dinesh et al. 2008b) and a complete revision with the description of twelve new species has been recently made by Biju et al. (2011) providing baseline for comparisons across taxa. The discovery of N. kumbara reiterates that there is need for systematic sampling across the Western Ghats for providing a comprehensive listing of amphibian species in the region. For delimiting species ranges and to appreciate mountain associated clade endemism in Nyctibatrachus , sampling has to be very systematic covering altitudinal, latitudinal and longitudinal gradients. Although N. kumbara sp. nov. is distinct from known species of Nyctibatrachus in morphology, acoustics, breeding behavior and single gene genetic study, we still warrant multiple gene phylogenetic studies on N. kumbara to substantiate its specific phylogenetic relationship with the other members of Nyctibatrachus . Apart from taxonomy and phylogeny, we stress the importance of studies on breeding behaviour, ecology and parental care in amphibians of the Western Ghats, which would provide a holistic and inclusive approach to understand the evolutionary significance of anurans in the Western Ghats.

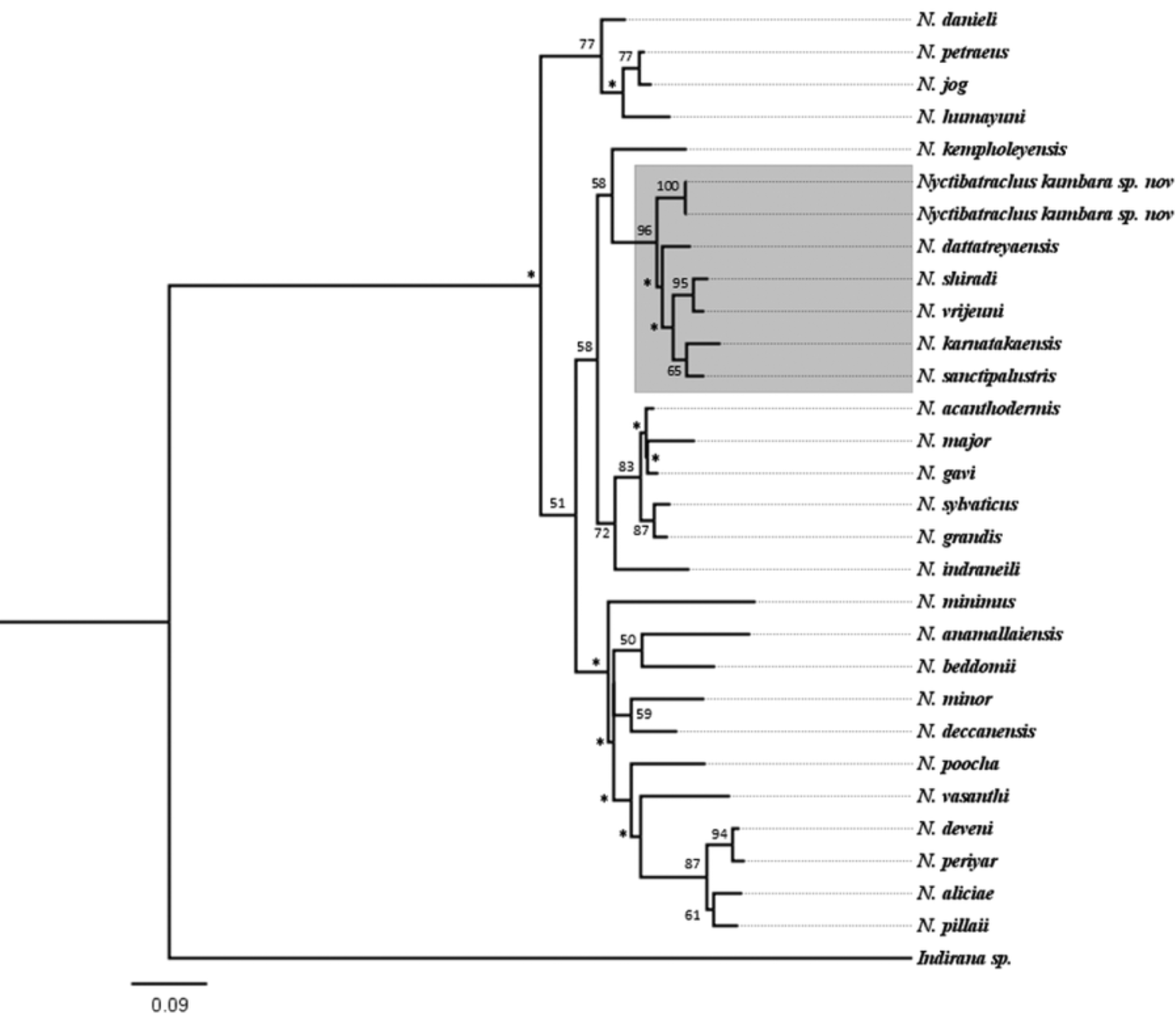







TABLE 5. Comparison of SVL (in mm, male and female) of allopatric and sympatric species of Nyctibatrachus using unpaired one tailed T-test with Bonferroni corrections for repeated measures. Numbers in parenthesis are p-values. Area in grey indicate allopatric species. NH _ M— Nyctibatrachus humayuni male, NH _ F— N. humayuni female, NP _ M— N. petraeus male. NP _ F— N. petraeus female, NJ _ M— N. jog male, NJ _ F— N. jog female, NK _ M— N. kempholeyensis male NK _ F— N. kempholeyensis female, NKU _ M— Nyctibatrachus kumbara sp. nov. male, NKU _ F— Nyctibatrachus kumbara sp. nov. female.
| NH_F n=8 | NP_M n=11 | NP_F n=8 | NJ_M n=13 | NJ_F n=9 | NK_M n=11 | NK_F n=9 | NKU_M n=6 | NKU_F n=5 | |
|---|---|---|---|---|---|---|---|---|---|
| NH_M n=9 | 1.53 (0.15) | 0.57 (0.58) | 0.27 (0.79) | 0.28 (0.78) | 0.1 (0.92) | 9.64 (<0.001) | 7.00 (<0.001) | 2.01 (0.07) | 1.86 (0.09) |
| NH_F | 1.25 (0.23) | 2.13 (0.06) | 2.25 (<0.05) | 2.07 (0.06) | 11.97 (<0.001) | 9.33 (<0.001) | 0.07 (0.95) | 0.14 (0.90) | |
| NP_M | 1.12 (0.28) | 1.23 (0.23) | 0.99 (0.34) | 15.11 (<0.001) | 11.54 (<0.001) | 1.95 (0.07) | 1.67 (0.13) | ||
| NP_F | 0.01 (0.99) | 0.27 (0.79) | 14.02 (<0.001) | 10.32 (<0.001) | 3.28 (<0.01) | 2.73 (0.02) | |||
| NJ_M | 0.3 (0.77) | 16.81 (<0.001) | 12.66 (<0.001) | 3.77 (0.002) | 2.97 (0.02) | ||||
| NJ_F | 17.3 (<0.001) | 13.15 (<0.001) | 3.49 (0.004) | 2.75 (0.03) | |||||
| NK_M | 7.44 (<0.001) | 22.19 (<0.001) | 15.49 (<0.001) | ||||||
| NK_F | 18.29 (<0.001) | 12.28 (<0.001) | |||||||
| NKU_M | 0.11 (0.91) |
TABLE 6. Size corrected pairwise comparison of call characteristic features of COCG species using ANCOVA. p-values in parenthesis. NJ _ pf / du— N. jog peak frequency and duration; NK _ pf 1 / du 1 — N. kempholeyensis peak frequency and duration for call Type I; NK _ pf 2 / du 2 — N. kempholeyensis peak frequency and duration for call Type II; NKU _ pf 1 / du 1 — Nyctibatrachus kumbara sp. nov. peak frequency and duration for single call; NKU _ pf 2 / du 2 — Nyctibatrachus kumbara sp. nov. peak frequency and duration for double call.
| NK_pf1/du1 | NK_pf2/du2 | NKU_pf1/du1 | NKU_pf2/du2 | |
|---|---|---|---|---|
| NJ_pf/du | 863.8 (<0.01) | 0.4 (0.54) | 0.2 (0.65) | 0.01 (0.97) |
| NK_pf1/du1 | 529.7 (<0.01) | 354.7 (<0.01) | 630.2 (<0.01) | |
| NK_pf2/du2 | 0.06 (0.82) | 0.28 (0.61) | ||
| NKU_pf1/du1 | 0.09 (0.77) | |||
| Discussion |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.