
Onychoquinpes permixtionis, Gómez & Morales-Serna, 2013
|
publication ID |
https://doi.org/ 10.1080/00222933.2012.757658 |
|
publication LSID |
lsid:zoobank.org:pub:2F77DCEA-37EE-428C-9E84-1F518F77512C |
|
persistent identifier |
https://treatment.plazi.org/id/C0DDD43D-C1B2-48CF-8526-941719E3ADA7 |
|
taxon LSID |
lsid:zoobank.org:act:C0DDD43D-C1B2-48CF-8526-941719E3ADA7 |
|
treatment provided by |
Carolina |
|
scientific name |
Onychoquinpes permixtionis |
| status |
sp. nov. |
Onychoquinpes permixtionis sp. nov.
( Figures 14 View Figure 14 –21)
Material examined
One female holotype (EMUCOP-080405-14), one male allotype (EMUCOP- 080405-15) preserved in alcohol, five dissected female (EMUCOP-080405-16, EMUCOP-080405-17, EMUCOP-080405-18, EMUCOP-080405-19, EMUCOP- 080405-20) and four dissected male paratypes (EMUCOP-080405-21, EMUCOP- 080405-22, EMUCOP-080405-23, EMUCOP-080405-24), and one CI, one CIV, and one CV paratypes (EMUCOP-080405-25), one male paratype (EMUCOP-080405- 26), one male and one female paratype (EMUCOP-080405-27), one female paratype (EMUCOP-080405-28), and four female and three male paratypes (EMUCOP- 080405-29) preserved in alcohol. Collected from El Verde estuary, Sinaloa, Mexico (23 ◦ 25 ′ 30 ′′ N, 106 ◦ 33 ′ 30 ′′ W), 8 April 2005, stns. 1 (sand, 3.38%, clay, 64.41%, silt, 32.21%; oxygen content, 2.7 ml l−1; salinity, 21 ‰; carbon content, 2.6%), 2 (sand, 5.8%, clay, 24.15%, silt, 80.52%; oxygen content, 2.7 ml l; salinity, 20 ‰; carbon content, 2.9%), 6 (sand, 6.46%, clay, 59.52%, silt, 34.01%; oxygen content, 3.7 ml l−1; salinity, 20 ‰; carbon content, 2.4%), 8 (sand, 10.71%, clay, 62.5%, silt, 26.79%; oxygen content, 5.6 ml l−1; salinity, 20 ‰; carbon content, 3.0%), coll F. E. Vargas Arriaga, F.N. Morales Serna and S. Gómez .
−1
Type locality
El Verde Estuary, Sinaloa State, north-western Mexico (23 ◦ 25 ′ 30 ′′ N, 106 ◦ 33 ′ 30 ′′ W).
Etymology
The specific epithet (Latin permixtionis , mixture) makes reference to the mixture of characters present both in Onychocamptus and Folioquinpes .
Descriptions
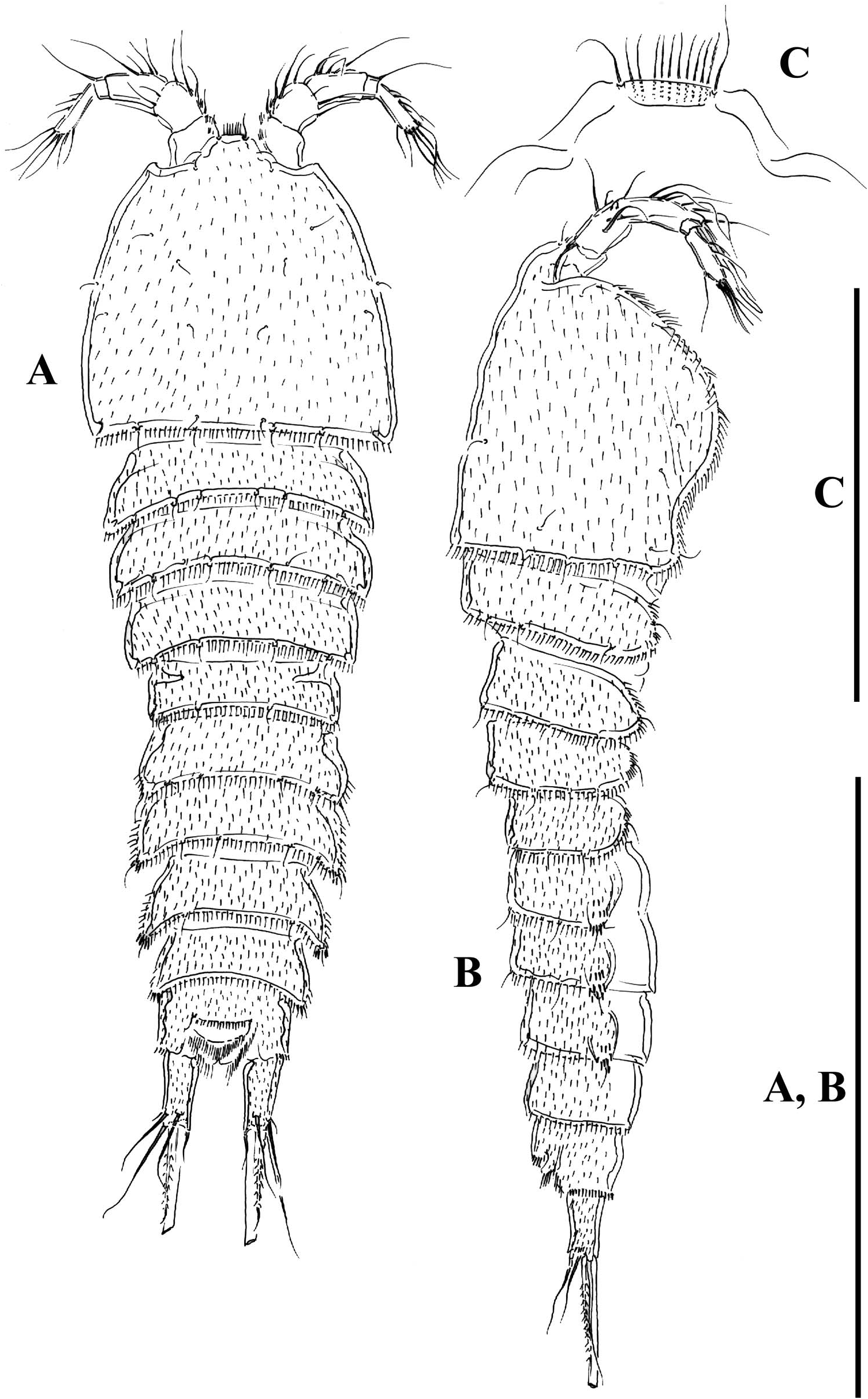
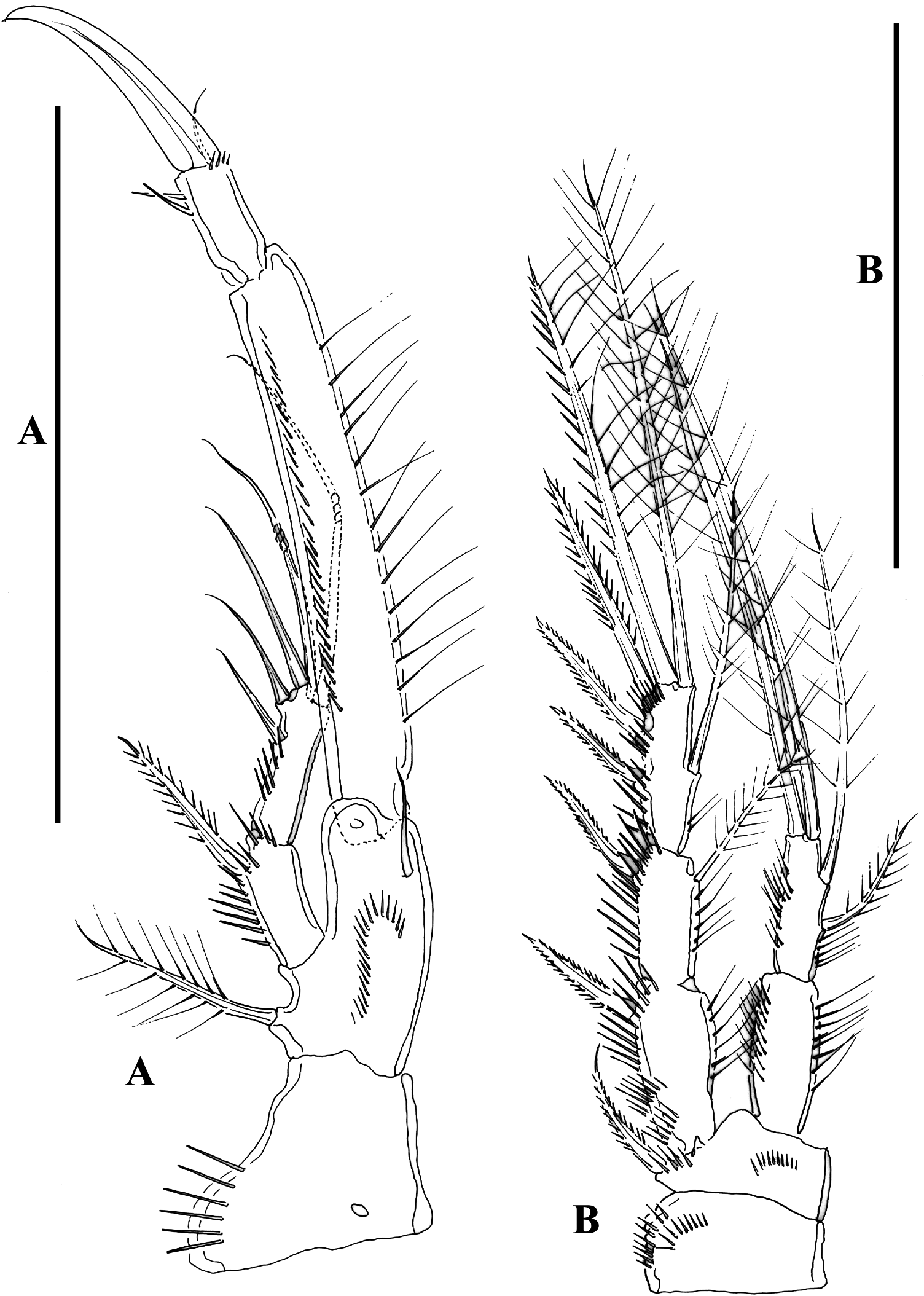
Female. Habitus ( Figure 13A, B View Figure 13 ), fusiform. Total body length measured from tip of rostrum to posterior margin of caudal rami, ranging from 405 µm to 535 µm (mean = 494 µm; n = 11). Cephalothorax, prosomites and urosomites covered with small spinules, and with row of small spinules along posterior and lateral margins. Rostrum ( Figure 13C View Figure 13 ) fused to cephalic shield, with flat tip, with row of setules ventrally, flanked by pair of sensilla. Genital-double somite distinct dorsally ( Figures 13A View Figure 13 , 14A View Figure 14 ) and laterally ( Figure 13B View Figure 13 ), fused ventrally ( Figure 14B View Figure 14 ); lateral processes of genital half moderately developed, with a sensillum arising from bulbous structure with tiny spinules; second half of genital double somite as previous somite dorsally, lateral processes moderately developed, with sensilla. Fourth urosomite as previous somite. Fifth urosomite as preceding somite, except for lack of sensilla in former. Anal somite ( Figure 14A, B View Figure 14 ) with ventral tube pores; posterior margin with spinules dorsally and ventrally; anal operculum rounded, posterior margin plain, flanked by two sensilla. Caudal rami ( Figure 14C, D View Figure 14 ) about 2.6 times as long as wide, with seven setae; seta I small, seta II dorsal to seta I, seta III visibly longer than seta II and situated laterally close to distal outer corner, seta IV nearly as long as seta II and fused basally to seta V, latter being longest, without fracture plan, seta VI small, arising at inner distal corner and about twice as long as seta I, seta VII situated dorsally close to posterior margin of ramus and biarticulated at base.
Antennule ( Figure 16A View Figure 16 ) five-segmented; first segment with medial and distal small inner spinules; first and second segments about 1.5 times as long as wide; third segment longest, about three times as long as wide; fourth segment smallest; fifth segment elongate. Armature formula, I-(1); II-(8); III-(9 + ae); IV-(1); V-(9 + (1 + ae)).
Antenna ( Figure 15A View Figure 15 ). Coxa as figured. Allobasis with one abexopodal seta, and with inner spinules. Exopod one-segmented, with two lateral and two distal elements, and with row of spinules. Free endopodal segment with inner row of spinules, with two spines and one slender seta laterally, and two strong spines, two geniculate single setae and one element fused to tiny seta basally.
Mandible ( Figure 16B, C View Figure 16 ). Strong gnathobase with bi- and multicuspidate teeth distally and one pinnate seta laterally. Palp small, one-segment, armed with one lateral and four apical elements.
Maxillule ( Figure 16D View Figure 16 ). Arthrite with five spines and a slender seta apically, and a slender seta laterally, with some spinules as depicted. Coxa with two setae. Basis with three elements (one element broken in Figure 16D View Figure 16 ). Exopod seemingly represented by one seta. Endopod represented by three elements.
Maxilla ( Figure 16E View Figure 16 ). Syncoxa with outer and inner spinules as depicted; with two endites, each with three setae. Allobasis drawn into strong claw with one anterior and one posterior accessory seta. Endopod represented by two long elements.
Maxilliped ( Figure 16F View Figure 16 ). Syncoxa with spinular rows as depicted, and with one apical seta; basis with small spinules along outer margin. Endopodal segment very small, with claw with one accompanying seta.
P1 ( Figure 17A View Figure 17 ). Coxa with several spinules as figured. Basis with longitudinal row of spinules, with naked inner element and pinnate outer element. Rami two-segmented. Exopod barely reaching proximal fifth of ENP1 because of the remarkable elongation of the basis at the insertion site of ENP; EXP1 with one outer spine; EXP2 with three spines and two geniculate setae. Endopod elongate; ENP1 about seven times as long as wide, ornamented as depicted; ENP2 about three times as long as wide, with some outer spinules, with one small apical seta and one strong claw, the latter about twice as long as supporting segment.
P2 ( Figure 17B View Figure 17 ). Coxa and basis ornamented as illustrated; the latter with outer pinnate spine-like element. Exopod three-segmented; EXP1 without, EXP2 with inner seta; EXP3 with three outer spines, two apical setae and one inner element. Endopod two-segmented, reaching slightly beyond EXP2; first segment without, second segment with two inner and two apical setae.
P3 ( Figure 18A View Figure 18 ). Coxa and basis ornamented as illustrated; the latter with long outer seta. Exopod as in P2. Endopod two-segmented, reaching tip of EXP2; first segment without, second segment with three inner, two apical, and one outer element.
P4 ( Figure 18B View Figure 18 ). Coxa and basis as in P3. Exopod as in P3 except for comparatively shorter EXP3. Endopod two-segmented, reaching half of EXP2; first segment without, second segment with one inner, one apical and one outer seta.
P5 ( Figure 15B View Figure 15 ) with separate rami. Baseoendopodal lobe well developed, with three setae plus outer seta of basis, and ornamented as shown. Exopod elongate, with two outer and one apical seta, the latter broad at its base, ornamented as depicted.
Armature formula as in Table 1.
Male. Habitus as in female, except for separate second and third urosomites ( Figure 18C View Figure 18 ). Total body length ranging from 325 µm to 350 µm measured from tip of rostrum to posterior margin of caudal rami (mean = 337 µm; n = 6).
Antennule ( Figure 19A, B View Figure 19 ) seven-segmented, subchirocer; fourth segment as in Figure 19B View Figure 19 ; fifth segment with two acute projections. Armature formula difficult to define: I-(1); II-(9);III-(8);IV-(10 + ae);V-(0);VI-(1); VII-(8 + (1 + ae)).
P1, P2 and P4 as in female; P3 EXP as in female; P3 ENP dimorphic ( Figure 19C View Figure 19 ), three-segmented, first segment as in female, second segment with inner seta and outer apophysis, third segment with two inner and two apical setae. P5 ( Figures 18C View Figure 18 , 19D View Figure 19 ) fused to supporting somite; ornamented as illustrated; with two setae (outer most smaller) plus outer seta of basis. P6 ( Figure 18C View Figure 18 , 19E View Figure 19 ) with spinules as figured, represented by two setae.
Remarks
The taxonomic history of the genus Onychocamptus is somewhat complicated and involves several events such as the establishment of the genus Echinolaophonte Nicholls ( Nicholls 1941) and further diagnosis of the genus Onychocamptus , creation of three subgroups within the genus ( mohammed -, horridus - and kliei -group) and reallocation of Nicholls’ (1941) species of Echinolaophonte in the horridus -group ( Lang 1948). Also, some species listed in Lang (1948: 1416) (i.e. Cleta minuticornis Buchholz , Laophonte hystrix Brian and Laophonte steueri Gurney ) were synonymized either with Laophonte horrida (= Echinolaophonte horrida ) or Laophonte armiger (= Echinolaophonte armiger ) ( Nicholls 1941; Lang 1948). Upon recognition of two forms of E. armiger, Lang (1965) suggested that this genus might be composed of two lines of evolution as evidenced by the armature formula of the swimming legs, and noted that, of the species referred to Onychocamptus , only the species of his mohammed -group remain (i.e. Onychocamptus mohammed (Blanchard and Richard) , Onychocamptus chathamensis (Sars) , Onychocamptus talipes (Wilson) and Onychocamptus bengalensis (Sewell)) .
Another part of this complex taxonomic history involves also the erection of the genus Klieonychocamptus Noodt ( Noodt 1958) to accommodate five species / subspecies ( Klieonychocamptus kliei (Monard) s. str., Klieonychocamptus kliei adriaticus (Petkovski) (comb. nov. for Onychocamptus kliei adriatica (Petkovski) (after Vervoort 1964)), Klieonychocamptus kliei confluens Nicholls , Klieonychocamptus diarticulatus Nicholls , and Klieonychocamptus discipes Noodt ). Serban and Pleşa (1957) described Klieonychocamptus ponticus (Serban and Pleşa) (comb. nov. for Onychocamptus ponticus Serban and Pleşa (after Vervoort 1964)), and Lang (1965) and Marcus (1971) relegated K. diarticulatus as a synonym of the former, and, in view of the great diversity in the male sexual dimorphism, Lang (1965) questioned the naturalness of Klieonychocamptus . This was also noted by Wells and Rao (1987), and Lee and Huys (1999) created the genus Psammoplatypus Lee and Huys to accommodate Paronychocamptus proprius Lang and K. discipes Noodt as Psammoplatypus proprius (Lang) and Psammoplatypus discipes (Noodt) , respectively.
Fiers (1998) demonstrated that O. talipes is a junior subjective synonym of O. mohammed , and Fiers and Rutledge (1990) created and diagnosed the genus Folioquinpes , with its type species, Folioquinpes mangalis Fiers and Rutledge being known from Papua New Guinea ( Fiers and Rutledge 1990), Louisiana ( USA) ( Fiers and Rutledge 1990; Rutledge and Fleeger 1993) and Malaysia ( Somerfield et al. 1998). In the same paper, Fiers and Rutledge (1990) removed O. chathamensis from Onychocamptus and placed the species into Folioquinpes as Folioquinpes chathamensis (Sars) . At present, F. chathamensis is known from the Chatham Islands ( Sars 1905), South Africa ( Ruhe 1914), India ( Sewell 1924), the Gulf of Mexico ( Suárez-Morales et al. 2009) and the Galapagos ( Mielke 1981). The establishment of Folioquinpes to accommodate F. mangalis and further reallocation of F. chathamensis was based primarily on the lack of sexual dimorphism of the male swimming legs of these two species and on the foliaceous female P5 EXP. Lee and Huys (1999) presented an updated diagnosis of Onychocamptus and considered the genus as an ancient lineage as evidenced by the retention of the inner proximal seta in the female and male P3 ENP2 (this primitive condition is present also in the cornuta -species group of Laophonte ( Lee and Huys 1999) , and in the subfamily Esolinae Huys and Lee , except for Troglophonte Huys and Lee where it is lost in both sexes ( Huys and Lee 2000)), and by the presence of an outer distal apophysis in the male P3 ENP2 (homologous to the outer element of the female P3 ENP2). Lee and Huys (1999) suggested that the lack of sexual dimorphism of the male swimming legs of Folioquinpes could be the result of heterochrony.
The new genus described herein exhibits a mixture of character states typical for Onychocamptus (presence of the primitive inner proximal seta in the female and male P3 ENP2, and the presence of an outer distal apophysis in the male P3 ENP2, and the armature formula of the P4 ENP2 (111)) and Folioquinpes (the foliaceous female P5 EXP, and the lack of sexual dimorphism in the male exopods) and is suggested to be placed in a new genus, Onychoquinpes gen. nov. The genus Folioquinpes bears a sister-group relationship with Onychocamptus ( Lee and Huys 1999) , which might also be the case for Onychoquinpes gen. nov. In their phylogenetic analysis, Huys and Lee (2000) confirmed the sister-group relationship between the Laophonte cornuta - group and Onychocamptus , and stressed that, even though the inclusion of advanced but related genera such as Folioquinpes might modify the relative position of Laophonte and Onychocamptus , the latter would show up as “an early speciation event predating the evolution of the other Laophontinae ” ( Huys and Lee 2000: 103) as evidenced by the retention of the ancestral setal formula of the male P3 ENP.
Further phylogenetic analysis in which genera such as Folioquinpes and Onychoquinpes gen. nov. are included, would confirm Lee and Huys’ (1999) hypothesis of a sister-group relationship between Onychocamptus , Folioquinpes and Onychoquinpes gen. nov. Hence, Onychocamptus would be basal to the other two genera with an inner proximal seta in the female and male P3 ENP2, and the presence of an outer distal apophysis on the male P3 ENP2 homologous to the outer element of the female P3 ENP2. Onychoquinpes gen. nov. would represent an early offshoot, sharing with Onychocamptus the primitive inner proximal seta on the female and male P3 ENP2, and the presence of an outer distal apophysis on the male P3 ENP2. Onychoquinpes gen. nov. and Folioquinpes share the general shape of the female P5 and the slight dimorphism in the armature of the male P3 EXP and P4 EXP. Within Folioquinpes , F. chathamensis seems to be the most primitive species sharing with Onychocamptus and Onychoquinpes gen. nov. the presence of a primitive inner proximal seta on the female and male P3 ENP2. Both species of Folioquinpes are unique by the lack of sexual dimorphism of the male P3 ENP2 (without outer apophysis). Folioquinpes mangalis seems to be the more advanced representative of this lineage given the lack of the inner proximal seta on the male and female P3 ENP2.





| CV |
Municipal Museum of Chungking |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |