
Pogonostoma (Pogonostoma) atrorotundatum W. Horn, 1934
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4881.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:DA5F7402-0D1E-42D7-9DBE-26DF54671F0C |
|
DOI |
https://doi.org/10.5281/zenodo.4328713 |
|
persistent identifier |
https://treatment.plazi.org/id/824987FA-DA55-FFD7-FF0E-FE5EFA95FCB0 |
|
treatment provided by |
Plazi |
|
scientific name |
Pogonostoma (Pogonostoma) atrorotundatum W. Horn, 1934 |
| status |
|
Pogonostoma (Pogonostoma) atrorotundatum W. Horn, 1934 View in CoL
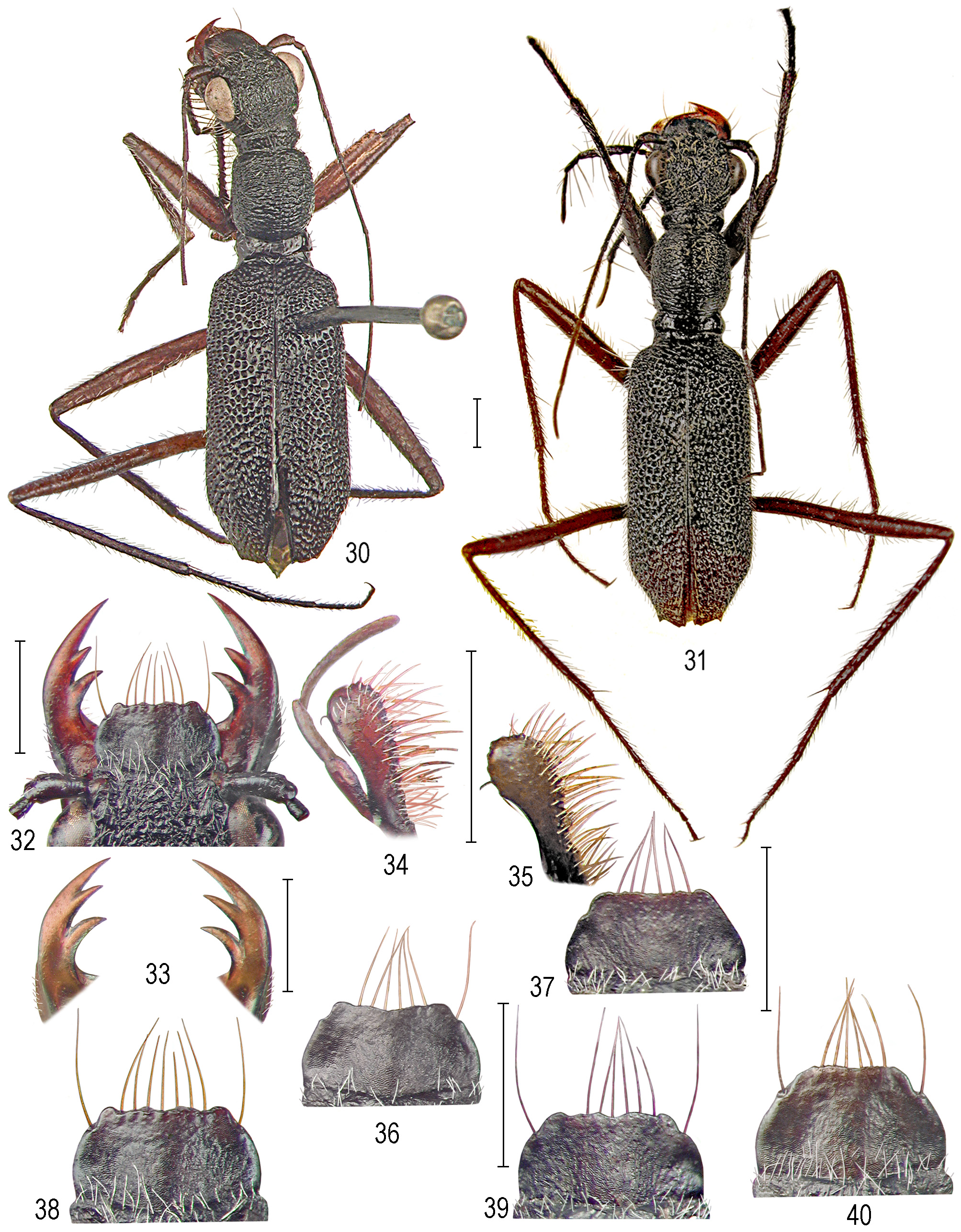
( Figs 30–66 View FIGURES 30–40 View FIGURES 41–49 View FIGURES 50–54 View FIGURES 55–66 , 105 View FIGURES 104–107 )
Pogonostoma elegans atro-rotundata W. Horn, 1934a: 28 and 34 [incorrect gender of the species-name].
Type locality. “Central-western Madagascar ” (see “ Distribution and biology” below) .
Pogonostoma (Pogonostoma) atrorotundatum: Jeannel 1946: 126 View in CoL (partim) [emendation of the species-name into neuter].
Pogonostoma (Pogonostoma) rufidens Rivalier, 1970: 299 View in CoL (synonymy by Moravec 2005: 22).
Type locality. “ Forêt de Bongolava ” (see “Distribution and biology” below) .
Pogonostoma (Pogonostoma) atrorotundatum: Moravec 2005: 50 View in CoL , figs 1–2, 7–17.
Pogonostoma (Pogonostoma) atrorotundatum: Moravec 2007: 269 View in CoL , figs 939–960, 1740–1742.
Misapplications.
Non Pogonostoma (P.) atrorotundatum sensu Jeannel (1946: 126 View in CoL , partim), which is partly P. (P.) rivalieri Moravec, 2005 View in CoL , nec P. (P.) atrorotundatum sensu Rivalier (1970: 299 View in CoL , 300, fig 13ar), which is P. (P.) rivalieri Moravec, 2005 View in CoL .
Type material of Pogonostoma elegans atrorotundatum W. Horn, 1934 . Lectotype (designated by Moravec 2005), ♁ in SDEI, labelled: “C. W Madag.” [handwritten] // “Type / W. Horn” [printed] // “ Syntypus ” [red, printed] // “Coll. W. Horn / DEI, Eberswalde” [printed] // “ Lectotype / Pogonostoma elegans / atrorotundatum W. Horn, 1934 / design. Jiří Moravec 2004 ” [red, printed] // “ Pogonostoma (s. str.) / atrorotundatum W. Horn, 1934 / det. J. Moravec 2004 ” [printed]. Paralectotypes. 1 ♁, 1 ♀ in SDEI: “C. W Madag.” [handwritten]. 2 ♀♀ in SDEI: “ Madagascar / Tananarive” [green, printed] // “Guy Babault” [printed] // “Mus. Paris” [handwritten/printed]. 1 ♀ in SDEI: “Tanan-arivo” [handwritten]. All paralectotypes in SDEI labelled: “Type / W. Horn” [printed] // “ Syntypus ” [red, printed] // “Coll. W. Horn / DEI, Eberswalde” [printed]. 1 ♀ in MNHN: “Museum Paris / Madagascar / Dr. R. Maire 1912” [green, printed] // “Clivius B...... [illegible] / Miandrivazo” [handwritten] // “Cotype / W. Horn” [printed]. All para-lectotypes labelled: “Revision J. Moravec 2003: / Paralectotype / Pogonostoma elegans / atrorotundatum W. Horn, 1934 ” [red, printed] // “ Pogonostoma (s. str.) / atrorotundatum W. Horn, 1934 / det. J. Moravec 2004 ” [printed].
Type material of syn. Pogonostoma (Pogonostoma) rufidens Rivalier, 1970 . Holotype ♁ in MNHN, labelled: “Ft. de Bongolava” [handwritten]; “rufidens / Rivalier / Type” [handwritten] // “Type” [red, printed] // “Museum Paris / J. Millot” [blue, printed]. Paratypes. 1 ♁ in MNHN with same locality label as in holotype. 6 ♁♁ in MNHN: “ Madagascar / Tananarive” [green, printed]. 1 ♁ in NHMW: “ Madagascar / Tananarive” [green, printed] // “ Pog. rufidens m. / Riv. det.” [handwritten] // “Coll. K. Mandl” [printed] // “Ideotype” (sic!) [red, printed]. 1 ♁ in CCJM (ex APCA) [its aedeagus extracted, mounted and stored separately by Rivalier]: “ Madagascar / Tananarive” [green, printed // “ Pog. rufidens m. / Riv. det.” [handwritten] // “ Pogonostoma (s. str.) / atrorotundatum W. Horn, 1934 / det. J. Moravec 2004 ” [printed].
Other material examined. 1 ♀ in MNHN: “ Ft. de Bongolava ” . 1 ♀ in MNHN: “ Tananarive ”. 1 ♁, 1 ♀ in CKWP: “ Madagascar / Antsalova , XII.1982 ” [blue, handwritten]. 1 ♁ in MNHN: “Museum Paris / Madagascar / col. Perrier de la Bathie, 1906” [green, printed]. 3 ♁♁ , 1 ♀ in CCJM, 1 ♁ in MHCW: “ Madagascar Ouest / Ant-salova, (Plateau du Bemaraha) / I.1978 A. Peyrieras ”. 1 ♁, 1 ♀ in CCJM: ibid. and: “collecteur loc.”. Recent data. 5 ♁♁ , 1 ♀ in JWCW, 5 ♁♁ in HSCA , 1 ♀ in CCJM: “ Madagascar / 16°05’S, 46°55’E / Mahajanga prov. / Ankara-fantsika Nat. Park, 6- 12.1.2002 / I. Andrew, V. Dolin & R. Andreeva leg.”. 2 ♁♁ in CCJM: “ Madagascar Ouest / Ambalatomby 16 km N de Boriziny ( Port Bergé ) / 30.I.2000, leg. Jiří Moravec ”. 3 ♁♁ , 2 ♀♀ in CJVB , 1 ♀ in CCJM: “ Madagascar, Sambirano / Ankaramy env. / Massif du Manongarivo / 3.II.2000, leg. Jan Vybíral ” .
Differential diagnosis. Pogonostoma (P.) atrorotundatum is distinguished from P. (P.) ondravybirali sp. nov. principally by its elongate aedeagus which in its left lateral view ( Figs 55, 57–60, 63, 65 View FIGURES 55–66 ) possesses only moderately bent basal half and notably elongate, more distinctly dorsad-bent rounded apex. Body dorsally black or with faint metallic lustre, rarely bright blue to violaceous-blue (see “Variability” and “Remarks” below). Mandibles generally darker with brownish-ferrugineous teeth or with only brownish apices of teeth (faded to testaceous-brownish in old specimens). For other differences and distinctions from others of the P. (P.) alluaudi species-complex see “Differential diagnosis” under P. (P.) ondravybirali sp. nov. above.
Redescription. Body ( Figs 30–31 View FIGURES 30–40 ) medium-sized, 9.00–11.5 (LT 10.0) mm long, 2.60–2.90 (LT 2.70) mm wide, black (faded to black-brown in old specimens) rarely with blue or violaceous lustre (faded or vanished in old specimens).
Head conspicuously narrower than body, width 2.00– 2.35 mm; temples very short, usually 2.8–3.4 times short-er than eyes).
Frons merging with clypeus in middle and not differentiated from vertex, moderately convex in middle; supraantennal keels consisting of elevated, mostly sharp anterior crest and lower and blunt posterior one.
Vertex with moderate posterior impression; frons-vertex surface irregularly scabrous-rugose, fine ridges on an-terior area acute, on median area vermicular and blunter, occipital area irregularly transverse-wavy rugulose; whole dorsal surface of head covered with white or blonde, decumbent and erect, hairlike setae.
Genae nearly smooth on anterior area, striate-rugulose on postgenal-temporal area, with sparse, thin setae.
Clypeus as in P. (P.) ondravybirali sp. nov. (and others of the species-complex).
Labrum with 4–7 anterior and 2 lateral testaceous setae; surface black (in old specimens fading to cinnamonbrown tinge in middle), shape generally as in P. (P.) ondravybirali sp. nov. (and others of the species-complex); male labrum ( Figs 36–39 View FIGURES 30–40 ) 0.55–0.70 mm long, 1.10–1.35 mm wide, female labrum ( Fig. 40 View FIGURES 30–40 ) longer, length 0.75– 0.85 mm, width 1.15–1.35 mm.
Maxillae ( Figs 34–35 View FIGURES 30–40 ): generally shaped, coloured and sized as in P. (P.) ondravybirali sp. nov. (in old specimens paler with cinnamon-brown tinge and lacinia with ochre-testaceous setae).
Palpi generally as in P. (P.) ondravybirali sp. nov.
Mandibles ( Figs 32–33 View FIGURES 30–40 ) shaped as in P. (P.) ondravybirali sp. nov. (and others of the species-complex), metallic-black with reddish-brown teeth and juxtamolar area or almost entirely ochre-brown to reddish-brown or mahogany-brown (faded to cinnamon-testaceous in old specimens).
Antennae in male longer than body, in female slightly shorter than body, scape black (pale-brownish in some old specimens), antennomeres 2–4 black (dark-brown in old specimens), 5–11 with greyish-rusty microtrichia (incomplete in type specimens).
Thorax. Pronotum ( Figs 50–54 View FIGURES 50–54 ) elongate, length 2.40–2.70 mm, width 1.65–1.85; anterior lobe slightly narrower than posterior lobe, its surface finely irregularly scabrous-rugulose and with scattered umbilicate tubercles bearing whitish hairlike setae; disc broadly ellipsoid in male, almost subglobose in female, notopleural sutures not obvious in dorsal view, median line indistinct, surface of disc coarsely irregularly transverse-rugulose; ridges anastomosing, short or elongate, or more continuous and transverse, usually more continuous in female, passing to mostly umbilicate tubercles on lateral areas (in both sexes); whole discal surface covered with scattered, long, decumbent or erect, hairlike, blonde to blackish setae (black setae better obvious on lateral areas); posterior lobe shiny, but with conspicuously uneven surface covered with large irregular foveae and setigerous punctures with blonde or blackened hairlike setae; lateral and ventral thoracic sterna as in P. (P.) ondravybirali sp. nov.
Elytra ( Figs 41–49 View FIGURES 41–49 ) oblong, rather stout 5.70–6.90 mm long, generally stouter than in P. (P.) ondravybirali sp. nov; elytral apex barely dimorphic; in male ( Figs 41–42, 45–48 View FIGURES 41–49 ), variably with small and blunt or distinct subacute or acute outer tooth, sinuously convex in middle (as in LT) or obliquely sloped towards small, usually blunt inner tooth, with very faint sutural emargination towards indistinct sutural spine; apex in female ( Figs 43–44, 49 View FIGURES 41–49 ) with blunt or right-angled external tooth, then variably rounded in middle or forming rather distinct but rounded inner tooth and moderately or very shallowly arcuately emarginated towards suture (sutural spine indistinct or undeveloped); elytral surface convex and with moderate discal impression, extremely coarsely punctate throughout except for sparser and finer punctures on basolateral area and smooth narrow basohumeral area (not obvious from above); punctures of nearly equal size, deep and very large with very thin, mostly sharpened intervals, occasionally anasto-mosing in irregular chains as often transversely opened with declined interval; punctures on apical area shallower and finer; elytral setal vesture consisting of nearly erect, copious, moderately long, whitish or dirtily blonde ornamental setae mixed with sparse, or more copious, black ones; sparse, long and erect, blonde, hairlike sensory setae are present particularly on humeral areas.
Legs. Coxae dark brown, covered with short, whitish setae; trochanters and femora concolorous with coxae, pro- and mesofemora pale-brown with white setae, metafemora almost black with dark setae; tibiae and tarsi blackbrown, protibiae and protarsi with whitish setae, mesotibiae and mesotarsi with mixture of white and black-brown setae, metatibiae and metatarsi with black-brown setae; hooks pale cinnamon-brown.
Aedeagus notably elongate, length 2.55–2.85 mm, width 0.30–0.40 mm., in its left lateral aspect ( Figs 55, 57–60 View FIGURES 55–66 ) with only moderately bent basal half and notably elongate, dorsally moderately bent and rounded apex; aedeagus in dorsal and ventral aspects ( Figs 56, 61, 62, 64 View FIGURES 55–66 ) constricted to narrow, cylindric apex.
Variability. The blue or blue-violaceous tinge is conspicuous on fresh specimens from northern areas (Ankarafantsika and Boriziny) but is not or only feebly obvious on old specimens including the type specimens (also those of synonymous P. (P.) rufidens ). Similarly, the black setae are rare and barely visible on elytra of most old specimens, but are usually more obvious on fresh adults from Ankarafantsika, and particularly on those from Boriziny. As this variability is seen on elytra of specimens with sympatric occurrence, the distinction was considered by the first author ( Moravec 2005, 2007) without taxonomic value. As also the mandibles of the adults from the abovementioned localities are prevailingly black except for reddish apices (both characters similar to P. (P.) alluaudi ), the adults from the above-mentioned areas may represent a further undesribed species of the species-complex. Notwithstanding, they possess the same characteristic shape of the aedeagus apex as in the lectotype and other specimens including those from south-western areas of Madagascar (for instance from Antsalova).
Distribution and biology. Pogonostoma (P.) atrorotundatum is a rather rare species, though distributed in a large area of central-western Madagascar and in Sambirano. As discussed previously ( Moravec 2007), the type locality “Central-western Madagascar ” was not exactly specified. As no specimen was designated as a type by Horn (1934a) or Olsoufieff (1934) and others until the syntype labelled “C. W Madag.” was designated as lectotype by Moravec (2005), the type locality was inaccurately mentioned as “Miandrivazo” by Jeannel (1946), followed by Rivalier (1970) who, however, clearly mentioned that the type is in “Museum Eberswalde” (now SDEI). As rectified previously ( Moravec 2005, 2007), Horn (1934a) mentioned in the original description of his Pogonostoma alluaudi atrorotundatum (as “atro-rotundata”): “Madag. centr. occident. et per errorem “Tananarive”, Miandrivazo et per errorem “Tananarive”. Collectio autoris et Museum Paris” It corresponds to the locality labels of the six syntypes (the male lectotype, one male and four female paralectotypes in SDEI and one female paralectotype (with “Miandrivazo” on its label) in MNHN cited in “Type material examined” above). Horn was quite right that the labels “Tananarive” or “Tananarivo” were erroneous as they were formerly synonyms for the whole island, when labelled as such (or as “Annanarivo”) by some historical insect dealers.
The type specimens of the synonymous P. (P.) rufidens Rivalier, 1970 are labelled “Ft. de Bongolava”. The very extensive massif of Bongolava stretches in the western area of central Madagascar, parallel with the Bemaraha plateau, and the type specimens of P. (P.) rufidens come from the region 75 km NW of Tsiroanomandidy, a forested stripe between Ambaravaranala and Beravina, 1250 m a.s.l. (A. Peyrieras, pers. comm., Viette 1991). Other non-type specimens, labelled “Antsalova”, come from the Bemaraha plateau (partly adjoining the Bongolava massif, which is the type locality of the synonymous P. (P.) rufidens ). Miandrivazo also lies in the Bemaraha plateau, southeast of Antsalova. It must be noted here that the above-mentioned forest of the Bongolava massif must not be confused with the homonymous, non-forested region “Bongolava” placed east of Majunga. The label “Mandritsara” probably means the area along the Sofia River in the montane area “Lembalemba Ambanin Androna”, 20–30 km north-west of Mandritsara in western Madagascar. The recently collected adults come from a deciduous forest in northern area of western Madagascar. Ankarafantsika National Park lies 25 km N of Ambato-Boeni.
In the locality Ambalatomby lying 16 km north of Boriziny (= Port Bergé), the adults were running along bark of large trunks (mostly of mango trees) in a secondary gallery-forest along the Sofia river, while in the forest of the Manongarivo massif (Sambirano) they preferred slim trunks of young trees.
Remarks. The five syntypes of P. elegans atrorotundatum in SDEI and one in MNHN examined by the first author ( Moravec 2005) well agree with the original description. Their aedeagi correspond with the original illustration of the aedeagus ( Horn 1934a, fig. 27a), and the shape is one of the main distinctive features of this species. The male lectotype was designated ( Moravec 2005) in order to avoid the confusion with P. (P.) rivalieri .
Two of the syntypes (paralectotypes) in SDEI are labelled “Mus. Paris”, but only one syntype is deposited in MNHN (the female from Miandrivazo with a white label “Cotype”). Jeannel (1946) illustrated a male from Miandrivazo which was not, however, found in the MNHN collection. Notwithstanding, the illustration of the aedeagus by Jeannel (1946, fig. 46e), though showing only an apical portion, does not correspond to the aedeagi of the syntypes of P. (P.) atrorotundatum and differs also from the above-mentioned original illustration by Horn (1934a, fig. 27a). As discussed previously ( Moravec 2007), in case that the aedeagus illustrated by Jeannel was drawn accurately, it was probably taken from a male from Antsirabe, instead from Miandrivazo mentioned by Jeannel, thus, probably from a male of P. (P.) rivalieri (specimens of both species were assembled in MNHN and SDEI un-der the name P. (P.) atrorotundatum ). Horn subsequently also confused these two different species, as obvious from their common arrangement in his collection (SDEI), and from the fact that one specimen (MNHN) from Antsirabe, identified by the first author as P. (P.) rivalieri , bears a label “ P. atrorotundatum W. Horn det. 1938”.
P. (P.) atrorotundatum sensu Rivalier (1970: 300 , Fig. 13 View FIGURES 11–16 ar) is a different species, described later as P. (P.) rivalieri Moravec, 2005 . Rivalier never examined the syntypes of P. atrorotundatum deposited in SDEI and did not take into consideration the original description or Horn’s illustrations ( Horn 1934a, pl. 1, fig. 27a, 1934b, pl. 1, fig. 27a) showing the distinctive shape of the aedeagus of the true P. (P.) atrorotundatum . The misinterpretation resulted in a superfluous description of P. (P.) rufidens Rivalier, 1970 . Some of the type specimens mentioned by Rivalier in the description of his new species (he mentioned only males as type specimens in the original descrip-tion), as well as the non-type females which were later identified by him as P. (P.) rufidens , bear the identical green labels “Tananarive” as the two paralectotypes of P. (P.) atrorotundatum in SDEI. Examination of type specimens in MNHN has disclosed ( Moravec 2005) that the characters of P. (P.) rufidens , including the characteristic shape of the aedeagus, well correspond to relevant diagnostic characters of the genuine type specimens of P. (P.) atrorotundatum W. Horn.
The two males in MRAC labelled “Mandritsara” listed previously ( Moravec 2005, 2007) as P. (P.) atrorotundatum must be examined again as they may represent another species of the P. (P.) alluaudi species-complex.



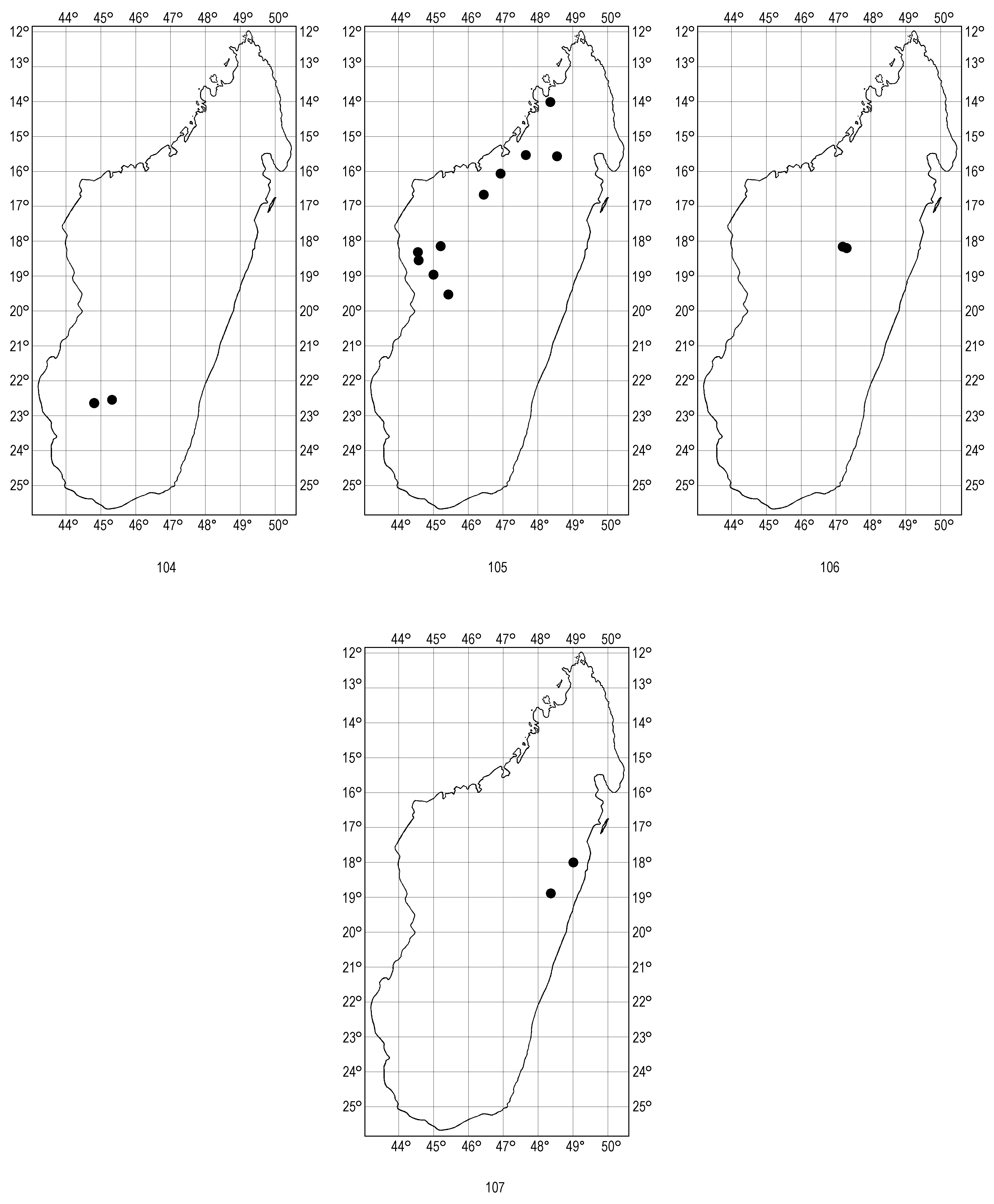

| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pogonostoma (Pogonostoma) atrorotundatum W. Horn, 1934
| Moravec, Jiří & Vybíral, Jan 2020 |
Pogonostoma (Pogonostoma) atrorotundatum:
| MORAVEC 2007: 269 |
Pogonostoma (Pogonostoma) atrorotundatum:
| MORAVEC, J. 2005: 50 |
Pogonostoma (Pogonostoma) rufidens Rivalier, 1970: 299
| MORAVEC, J. 2005: 22 |
| Rivalier, E. 1970: 299 |
Pogonostoma (Pogonostoma) atrorotundatum:
| Jeannel, R. 1946: 126 |
Pogonostoma (P.) atrorotundatum sensu
| Rivalier, E. 1970: 299 |
| Jeannel, R. 1946: 126 |
Pogonostoma elegans atro-rotundata W. Horn, 1934a: 28
| Horn, W. 1934: 28 |