
Rhampholeon (Rhinodigitum) hattinghi, Tilbury, Colin R. & Tolley, Krystal A., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3905.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:5AFBC257-AB6C-4DB0-8A43-97B604F77DEC |
|
DOI |
https://doi.org/10.5281/zenodo.6097647 |
|
persistent identifier |
https://treatment.plazi.org/id/03A63820-7932-FF84-FF24-56B4FC7A0FA6 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhampholeon (Rhinodigitum) hattinghi |
| status |
sp. nov. |
Rhampholeon (Rhinodigitum) hattinghi sp. nov.
Synonymy: Rhampholeon boulengeri Tilbury 2010 .
Holotype. PEM-R19194, an adult male with everted hemipenes, collected by Colin Tilbury and Isak Hattingh on 21st March 2010, from path side vegetation at 1700 metres a.s.l., Mount Nzawa, Moba District, Katanga Province, Democratic Republic of the Congo (6° 51.07’ S; 29° 35.87’ E).
Paratypes. PEM-R 19193, adult male; PEM-R19195, adult male, PEM-R19196, adult female; PEM-R19197, adult female; PEM-R19198, adult female; all collected on the same date and locality as the holotype.
Diagnosis. Due to its markedly bicuspid claws, amelanotic parietal peritoneum and acalyculate bag-like hemipenes with dual apical horns, this taxon can be placed within the sub-genus Rhampholeon (Rhinodigitum) Matthee et al. 2004 . In spite of the remarkably conserved external morphology within this genus, this new taxon is easily distinguishable from congeners by virtue of the following identifying characteristics: The consistent absence of deep mite pockets (or pits) in the inguinal flexure distinguishes this taxon from Rh. beraduccii Mariaux & Tilbury and the six species of the Rh. platyceps Günther complex ( Branch et al. 2014), including Rh. platyceps , Rh. chapmanorum Tilbury , Rh. maspictus Branch et al. , Rh. tilburyi Branch et al. , Rh. bruessoworum Branch et al. and Rh. nebulauctor Branch et al. ; the presence of deep pits in the axillae of this taxon distinguishes it from Rh. nchisiensis Loveridge and Rh. acuminatus Mariaux & Tilbury ; the relatively smooth supra-orbital and canthal crests distinguish this taxon from Rh. boulengeri , Rh. uluguruensis Tilbury & Emmrich and Rh. moyeri Menegon et al. , in all of which a peaked cluster of tubercles forms distinct protuberances above the eye and the nasal aperture ( Fig. 5 View FIGURE 5 ).
Description of Holotype. Snout-vent 56mm, tail 11mm. Body habitus leaf like - typical of all other Rhampholeon (Rhinodigitum) species. Head short, casque flattened, top of head shallowly concave. The lateral crests are studded with several prominent tubercles. Parietal crest indistinct, indicated by a short row of 3 marginally enlarged tubercles. The supra-optic ridge gathers into a low cluster of tubercles anteriorly above each eye, but without forming a supra-optic horn. The two supra-orbital ridges are connected to each other by a series of 18 inter-orbital tubercles arranged in a shallow V across the top of the head. The canthal ridges are formed by a row of enlarged, relatively smooth tubercles which terminate anteriorly at the base of a small, finely tuberculated stump-like process that barely projects off the anterior end of the snout. The nares open posteriorly, from a small bulge at a point roughly one third of the distance between the anterior orbital rim and the front of the maxilla. A distinct temporal crest arises from the mid post-orbital rim and consists of 5 tubercles on the right and six tubercles on the left, of which the most posterior is the largest. Three large sub-conical tubercles are spaced along the inferoposterior rim of the orbit. A prominent tubercle is situated just above the mouth in line with the posterior orbital rim.
The dorsal vertebral line is weakly crenulated, fading to smooth over the pelvic region. The dorsal margin of the tail is likewise smooth.
Deep axillary mite pockets (or pits) are present, whilst the inguinal flexures are smooth.
The background scalation of the body, tail, limbs and belly is composed of tightly packed sub-homogeneous, stellate-edged tubercles. The gular region of the throat extending to between the front limbs is studded with many small conical tubercles. These also are found on the belly, underside of the tail and the underside and particularly on the outer-surface of the limbs. The sides of the body have regularly-spaced, enlarged conical tubercles, with three particularly enlarged cones situated in a row at midbody, the first sited above the shoulder, the second at midbody, and the last just before the pelvis.
Claws are markedly biscuspid, several prominent palmar and plantar accessory spines are seen at the base of the toes, and the outer soles of the feet are relatively spinose whilst the inner soles are smooth with a cobblestone appearance.
Colour in life. Adult male ( Figs 6 View FIGURE 6 & 7 View FIGURE 7 ). Top of head and snout dark brown extending to cover the sides of the snout above the line of the mouth and to a lesser extent the eyeball and superior temporal zone. Eyeball with dark radiations especially prominent over the upper half of the eyeball. Gular region off-white. Body light brown mottled with olive green. Five dark blotches are spaced along the dorsal vertebral ridge, and a broad dark blotch is also located over the dorsum of the tail. Two thin dark green stripes, originating from the second and third vertebral blotches, extend infero-posteriorly over the flanks. Three dark spots are arrayed along the flank, each spot centred over a prominent conical tubercle. The inner sides of the fore and hind limbs pale orange. A prominent white spot is located on each side of the tail.
Adult female stress pattern ( Fig. 8 View FIGURE 8 ). Dorsal ridge orange from the tip of the casque to the tip of the tail. Flanks speckled with small orange spots and three large white spots on the flanks, each surround an enlarged conical tubercle. Sides of head and gular region flecked in white, sometimes forming short vertical stripes. Eyeball dark and speckled with light blue tubercles. Lateral flank stripes dark but heavily flecked with light blue.
Hemipenes. Short bag-like, truncus acalyculate; a pair of short, broad-based apical horns incurving above the sulcus spermaticus ( Fig. 9 View FIGURE 9 ). The upper surface of the horns is covered in closely packed thorn-like papillae, much like a pineapple skin. Fourteen papillae are arranged in a proximal or basal cluster terminating in a distal row of two papillae. Inferior to each apical horn, a prominent balloon-like membranous evagination billows anteriorly.
Sexual Dimorphism. Apart from the prominent hemipenal bulges there is little sexual dimorphism evident in the type series ( Table 4 View TABLE 4 ). In males the tail comprises between 16–23% of the total length whilst in females it is between 14–18% of total length.
Number Sex Length Rp Casque/snout Sub-orbital Axil pit Inguinal pit
(mm) (mm) rows
*= Holotype, measurements in millimetres; s-v = snout-vent length; Io tub = inter-orbital tubercles; Io diam = interorbital diameter; Length Rp = length of rostral process; Sub-orbital rows = number of rows of tubercles between the inferior orbital rim and the upper labials; na = not measured.
Variation. In the paratypes the parietal crest was either absent, or merely indicated as in the holotype. The supra-orbital crest is relatively smooth in all specimens, only forming a low cluster of tubercles in 2 specimens at the point where the inter-orbital tubercles terminate above the eye. The soles of the feet may vary from relatively smooth to a sub-acuminate field of tubercles. The hemipenes of PEM-R19193 and PEM-R19195 do not differ in any substantial way from that of the holotype.
Reproduction. The parietal peritoneum in the abdominal cavity of the largest female (PEM-R19198) was unpigmented and the fallopian tubes each held a single egg measuring 6.4mm x 5.6mm.
Habitat. All specimens were found in closed canopy Afrotemperate montane forest on low vegetation alongside a path. Perch heights varied from a few centimetres up to 50 cm from the ground.
Etymology. Named for Isak Hattingh, who enthusiastically but unwittingly agreed to accompany the first author on a trip which tested many boundaries. He never complained once.




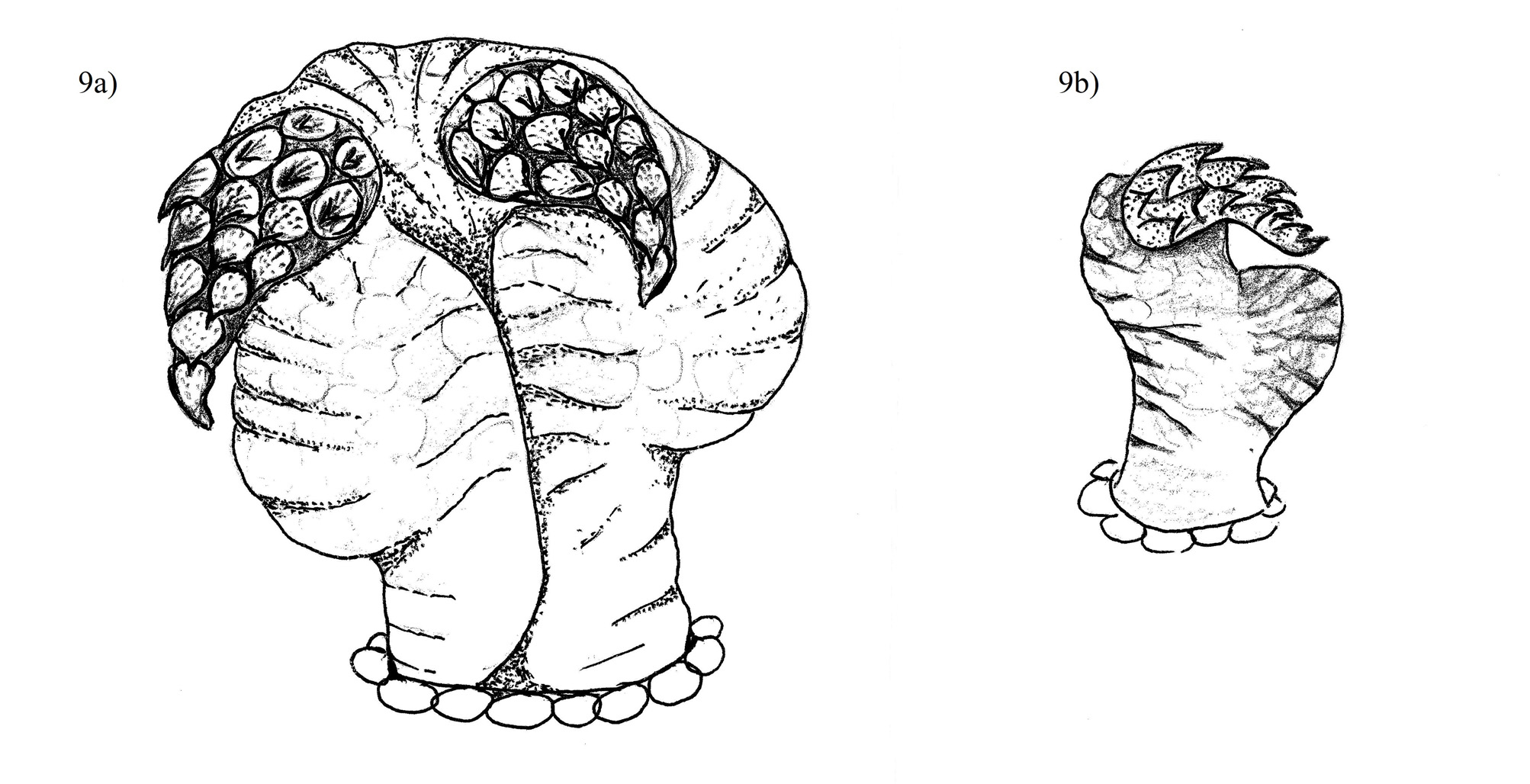
TABLE 4. Morphometric measurements for the type series of Rhampholeon hattinghi. sp. nov.
| Number | Sex | s-v (mm) | Tail (mm) | % t/total length Io tub | Io diam |
|---|---|---|---|---|---|
| PEM-R 19193 | Adult male | 47 | 12 | 20 17 | 5.8 |
| PEM-R 19194 * | Adult male | 56 | 11 | 16 18 | 6.54 |
| PEM-R 19195 | Adult male | 44 | 13 | 23 17 | 5.9 |
| PEM-R 19196 | Adult female | 50 | 11 | 18 | 6.23 |
| PEM-R 19197 | Adult female | 45 | 10 | 18 15 | 5.5 |
| PEM-R 19198 | Adult female | 56 | 9 | 14 18 | 6.0 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.