
Scolopendra longipes Wood, 1862 Revalidated
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4111.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:4F24848F-2205-45F2-BC6C-AD56A00E153E |
|
DOI |
https://doi.org/10.5281/zenodo.5674267 |
|
persistent identifier |
https://treatment.plazi.org/id/1F52879D-C361-FFE0-93AC-FA92FB07FE5C |
|
treatment provided by |
Plazi |
|
scientific name |
Scolopendra longipes Wood, 1862 Revalidated |
| status |
|
Scolopendra longipes Wood, 1862 Revalidated View in CoL
( Figs. 1 View FIGURE 1 A, 2A, D, 3, 4 A–B, 5A, 6 A–B, 7 A–B, 8; Table 1 View TABLE 1 )
Scolopendra longipes View in CoL — Wood, 1862: 26 (nec S View in CoL . alternans Pocock 1893, nec S View in CoL . alternans Kraepelin 1903, nec S View in CoL . alternans Chamberlin 1914, nec S View in CoL . alternans Attems 1930, nec S View in CoL . alternans Shelley 2002, nec S View in CoL . alternans Mercurio 2010).
Type locality. The original type locality should be: USA: Florida, Ft. Jefferson, Garden Key, Dry Tortugas, received by Wood from Dr. D.D. Whitehouse, measuring 115 mm; however, the type material for S. longipes cannot be found at the USNM despite the fact that Wood states all of the listed specimens belonged to the Smithsonian Institution. After reading Roth’s (1959) biographical memoir on Wood it was discovered that while Wood’s original manuscript of The Myriapoda of North America was waiting for publication it was destroyed by a fire on January 24, 1865 at the Smithsonian. Unfortunately, it is highly likely that the type material for S. longipes was also destroyed during this fire. As far as can be ascertained, no original type material exists, therefore I designate the following type: NEOTYPE: Unknown sex from Florida, Monroe Co., Sugar Loaf Key, 2.7 miles S. of junction U.S. RTE 1 and FLA 939 (at sugar loaf channel, on NE end of island) on FLA 939, 24°37ʹ28.3ʺN 81°31ʹ06.6ʺW, 15 March 1977, 1400 hrs., sunny, 86°F, S. Blair Hedges, col., Under moist cardboard lying on the ground in the shade of a mangrove tree on the Atlantic side of Rte. 939 (1, USNM), measures 98 mm. At some point a topotype should be obtained and genetically compared to a specimen from this neotype location to assure that this topotype specimen is not a subspecies.
Material examined. Bahama Islands: N. Bimini, Feb. 17, 1970, V. Roth, (1, AMNH); South Bimini, May, 1951, W.J. Gertsch & M.A. Cazier, (2, AMNH); South Bimini, May, 1951, Gertsch & Cazier, (2, AMNH); So. Bimini, No. Shore, Jan. 1, 1952, (1, AMNH); west end of South Bimini Island, under rotten palm log, Sept. 26, 1947, James Oliver, (1, AMNH); Bimini, Oct. 6, 1947, James Oliver, (1, AMNH); Nassau, New Providence, May 31, 1904, W.W. Wheeler (A3245, Cat. No. 1542), (1, AMNH); Nassau, New Providence, West Bay, May 29, 1904, W.W. Wheeler (A3246, Cat. No. 1541), (1, AMNH). Bimini, obtained from GHF in 1998 (1, RJM). USA: Florida, Key Largo, Feb. 25, 1906, S.A. Binion (A3339, Cat. No. 1563), (1, AMNH); Monroe Co., Boot Key, April 1, 1996, R. Lawson & P.G. Frank (2, CAS); Miami, July, 1918 Mus. Exped., W.I. Charlesworth col., Rec’d from Dept. of Botany, Aug. 15, 1922 (3, FMNH); Monroe Co., Plantation Key, 4 mi. S. of Tavernier, 11.III.1963, H.L. Levi, edge hardwood forest, (1, MCZ); Monroe Co., Little Torch Key: L9/ 15, 28.i. 79, Coll. J. Lazell (1, MCZ); Monroe Co.; No Name Key; 1.1 miles E., 0.25 miles S. Bogie Channel Bridge, 16 Jan 1978, 0 930 hrs., ca. 57°F, under trash, S. Blair Hedges (1, USNM); Monroe Co., No Name Key, 1.5 miles E. Bogie Channel Bridge, 16 Jan 1978, 1000 hrs., ca. 60°F, under cardboard, S. Blair Hedges (1, USNM); Monroe Co.; Largo Key, ca. 6.3 miles NNE Key Largo (2.5 miles N. jct. RTE 1 on FLA. 905) 14 January 1978, 1730 hrs., ca. 62°F, under trash on east side of RTE 905, S. Blair Hedges (1, USNM); Chapman Field, Sept. 1928 (1, USNM); Monroe Co., Sugar Loaf Key, 2.7 miles S. of junction U.S. RTE 1 and FLA 939 (at sugar loaf channel, on NE end of island) on FLA 939, 15 March 1977, 1400 hrs., sunny, 86°F, S. Blair Hedges, col., Under moist cardboard lying on the ground in the shade of a mangrove tree on the Atlantic side of Rte. 939 (1, USNM); Florida, obtained from SI on November 6, 2012 (2, RJM); Florida, obtained from SI on November 7, 2013 (6, RJM).
Redescription. This redescription is based on live and preserved specimens observed, and is not drawn from a single specimen. Length up to 152 mm. Figure 1 View FIGURE 1 A illustrates the overall body shape and coloration in life. The cephalic plate, 1st tergite, forcipular segment and tergites 20–21 are burnt sienna. The burnt sienna color may continue further towards the middle of the trunk from 1st tergite and 20th tergite, but the midsection of the trunk generally appears lighter varying from russet to tawny. In recently preserved specimens (70–75% ethanol), the trunk tends to blend into a uniform tawny while the head and forcipular segment remain a burnt sienna.
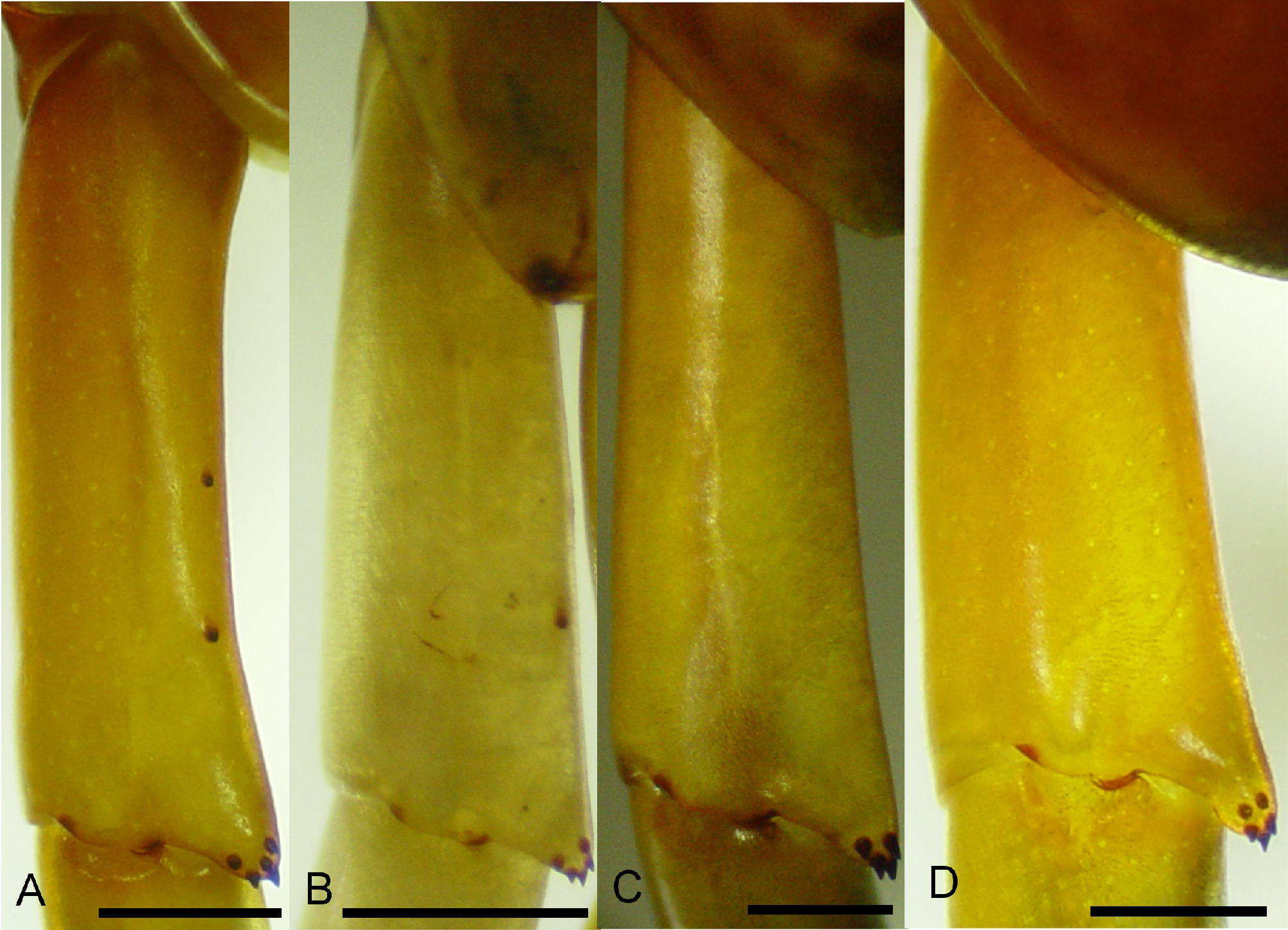
Antennal segments 16–18, with 17 being the most common. The first five antennal segments are glabrous with the remaining distal segments being hirsute. The cephalic plate is slightly longer than wide with a median ratio of 1.06 and a range of 1.01–1.09. The lateral margins of the head gently converge anteriorly, posterior edge rounded, paramedian sutures complete and diverging anteriorly, and sparsely punctate (~3–4/mm2) with punctation due to sensillia trichodea. These sensillia trichodea are very conspicuous over the entire body ( Figure 3 View FIGURE 3 ). The three medial teeth of coxosternal tooth-plate are fused, with or without pronounced invagination between the second and third teeth ( Figures 4 View FIGURE 4 A–B). The forcipular tergite lacks an anterior transverse suture but has complete paramedian sutures and these continue through tergite 20. Excluding the questionably large specimens, the maximum tergite width was 9.2 mm, which occurred on the 10th tergite. The ultimate tergite lacks a medial longitudinal suture ( Figure 5 View FIGURE 5 A). Sternite 2 has incomplete paramedian sutures beginning anteriorly and appear to end approximately 1/ 3 the sternite length with very faint continuation, if any, posteriorly. Sternites 3–20 have complete paramedian sutures and appear more pronounced posteriorly. The posterior edge of the ultimate sternite is gently rounded to rounded with the latter seemingly more common in smaller specimens ( Figures 6 View FIGURE 6 A–B). Tarsus one of leg one with a dorsal anterior spur and legs 1–20 each with a ventrodistal spur. The eighteenth prefemur of left or right leg may rarely have a single spine on the distomedial process. The antepenultimate prefemur usually has two spines on the distomedial process; but sometimes only 1 may be present. The penultimate prefemur has 4–6 spines on the distomedial process and typically 1–2 spines on dorsomedial surface ( Figures. 7 View FIGURE 7 A–B). The dorsomedial spines of the penultimate prefemur on younger specimens may vary slightly from what is seen in most specimens, such as two on left and none on right. Older specimens may show an additional spine on left or right, such as three on left and two on right. The ultimate prefemoral process typically has 6–8 spines with 7 being the most common. The maximum length to maximum width of the ultimate prefemur has a median ratio of 3.75, with a range of 3.65–3.95 ( Figure 8 View FIGURE 8 ). The total spine count on the ventral, medial, and dorsal surfaces of the ultimate prefemur ranges from 28–37, excluding the distomedial process. The coxopleural pores are either uniformly minute with a smooth appearance, as in S. alternans ( Figures. 6 View FIGURE 6 C–D), to a mixture of small and minute pores giving a rough appearance ( Figures. 6 View FIGURE 6 A–B). The lateral coxopleural spines are typically two, with three on occasion; and the third being closer to the coxopleural process, which typically has 7–10 spines.
Geographical distribution. From Dry Tortugas through Florida Keys and southern Florida including the counties of Monroe, Dade and Collier, to the Bahamas.
Etymology. The specific name longipes means long-foot and refers to the long, thin ultimate legs found in S. longipes .
Remarks. Evidently the description of S. longipes by Wood has never been thoroughly reviewed by subsequent authors, because there is one difficult-to-see character he mentions that clearly differentiates S. longipes from S. alternans . In Wood’s (1865) English description he clearly states, “…basal joint of penultimate pair armed with 5 spines on a terminal angular process and 1–2 others;” Wood’s “basal joint” actually refers to the penultimate prefemur. Therefore, the key differentiating characters here are the 1 or 2 other spines, because his language strongly suggests he was not indicating 1 or 2 additional spines on the prefemoral process but was actually referring to the spines found on the dorsomedial surface of the penultimate prefemur.
Wood (1862, 1865) mentions three characters of S. longipes that differ from Newport’s (1844, 1845) characters of S. alternans . Firstly, the process of the forcipular trochanteroprefemur (which Wood called the “mandibular tubercle” and Newport referred to as the “mandibular tooth.”) was claimed by Wood to be “…very large and having the lesser tubercle near to its base…” and Newport described the “…mandibles strong, with the tooth small, but armed with a minute tubercle near its apex…” The process of the trochanteroprefemur in S. longipes and S. alternans appears very similar and when taking variability into account does not provide good characters to distinguish these two species. Secondly, Wood stated that the number of spines on the prefemur of the ultimate pair of legs was 30–45 in S. longipes but 45–60 in S. alternans . Specimens of S. alternans with anywhere near a total of 60 spines on the ultimate prefemur were not observed in this study. Newport was very clear in stating that 30–40 spines were on the dorsal and medial surfaces while 15–20 were on the ventral surface. The maximum number of spines seen on the dorsal and medial surfaces combined in S. alternans is 21 in addition to 17 on the ventral, but this is another character that is fraught with difficulty because it is frequently hard to determine if a spine should be labeled as dorsal/medial or medial/ventral. Perhaps a standardized method of describing the positions of the ultimate prefemur spines can be derived, but at the moment a foolproof way of assessing these spines in the S. alternans -complex is elusive. Newport said the locality for his specimens of S. alternans was “Caribbean Islands,” which is highly indefinite, but this difference in spines on the ultimate prefemur suggests he may not have been looking at a specimen from Florida, Cuba, Haiti or Puerto Rico. Therefore, the 40–60 spines on the ultimate prefemur of S. alternans as opposed to the 30–45 in S. longipes cannot be corroborated as a good character to separate these species. Finally, referring to the ultimate coxopleuron of S. longipes, Wood mentions the roughness of the “lateral anal appendages”. He was surely referring to the ultimate coxopleura of S. longipes , which can have an evenly distributed pattern of variably sized, small to minute coxal pores ( Figures 6 View FIGURE 6 A, B) giving a rougher look than those of S. alternans , with uniformly sized minute coxal pores, evenly distributed, and a smoother appearance ( Figure 6 View FIGURE 6 C, D). It should be noted that specimens of S. longipes from various Florida Keys have uniformly minute pores giving a rather smooth appearance, similar to what would be considered S. alternans . This coxopleural pore variability, along with other variable characters and extended periods of isolation on relatively small islands, suggests that S. longipes may also actually be composed of subspecies. Incidentally, the diameter and surface area density of these coxopleural pores may be useful distinguishing characters, as well as that of the sensillia trichodea.
The range in length of the five specimens of S. longipes that Wood (1862) reported was 95.25–127 mm. In living specimens of S. longipes , the maximum adult length and weight ranges observed in this study were 105–128 mm and 5.3–6.4 g respectively. The longest specimen was from Chapman Field, Miami, which was about 152 mm but was poorly preserved.
During the course of this study, three of the live S. longipes laid eggs and brooded between 30–43 offspring. Although most of them were raised, some didn’t survive and were preserved. After examining four offspring from one female, only one specimen had a single faint spine on the right dorsomedial surface of the penultimate prefemur. This indicates that these spines are added with age, so they may not typically be present on specimens that are ≤ 30 mm long. Two of the juvenile specimens that were well preserved had an ultimate prefemur length to width ratio of 3.82, which was consistent with that of the adults. The Bimini Island specimen is young and has 1 dorsomedial spine on each penultimate prefemur ( Figure 7 View FIGURE 7 B). The two smaller live specimens of S. longipes from Florida (~ 100 mm), which are a little larger than the Bimini specimen (~ 75 mm), also have one dorsomedial spine on both penultimate prefemurs. Based on the material examined in this study, the addition of the second dorsomedial spine (distal) on each penultimate prefemur of S. longipes appears to occur with age.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Scolopendra longipes Wood, 1862 Revalidated
| Mercurio, Randy J. 2016 |
S
| Mercurio 2010 |
S
| Shelley 2002 |
S
| Attems 1930 |
S
| Chamberlin 1914 |
S
| Kraepelin 1903 |
S
| Pocock 1893 |