
Vermetus (Thylaeodus) enderi Schiaparelli, 2000
|
publication ID |
https://doi.org/ 10.5281/zenodo.5394546 |
|
publication LSID |
lsid:zoobank.org:pub:DBB169D6-A0CC-48E5-AE11-7C6201CC943C |
|
persistent identifier |
https://treatment.plazi.org/id/AD2A87E5-FFF5-ED1F-5B62-1699FB3BFA09 |
|
treatment provided by |
Marcus |
|
scientific name |
Vermetus (Thylaeodus) enderi Schiaparelli |
| status |
sp. nov. |
Vermetus (Thylaeodus) enderi Schiaparelli View in CoL n. sp.
Serpulorbis roussaei Vaillant, 1871 . – Springsteen & Leobrera 1986: 44, pl. 8, fig. 10 (not Vaillant 1871).
Serpulorbis (s.s.) roussaei (Vincent) (sic) – Savazzi 1996: 160, fig. 2A (not Vaillant 1871).
Serpulorbis roussaei of authors (not Vaillant 1871).
TYPE MATERIAL. — Holotype and paratypes: in an aggregate of specimens ( MNHN) ( Fig. 2C View FIG ) ; paratypes: one aggregate of specimens ( MZB 14024) ( Fig. 2B View FIG ), one aggregate of specimens ( MSNG) ( Fig. 2A View FIG ), one aggregate of specimens on Lopha sp. , coll. Schiaparelli. TYPE LOCALITY. — Kudarah Thila ( Shoal of Kudarah ), 72°55’E, 3°34’N, South Kudarah Island , Ari Atoll, Maldives, Indian Ocean GoogleMaps .
ETYMOLOGY. — From the Divehi (Maldivian language) name ( ènderi ) of the black coral ( Antipathes spp. ), on which this species is commonly found.
MATERIAL EXAMINED. — Several specimens from type locality, Philippines and Indonesia.
KNOWN DISTRIBUTION. — Maldives, Philippine Islands (Cebu) and Indonesia (Bunaken Marine Park, North Sulawesi).
DESCRIPTION
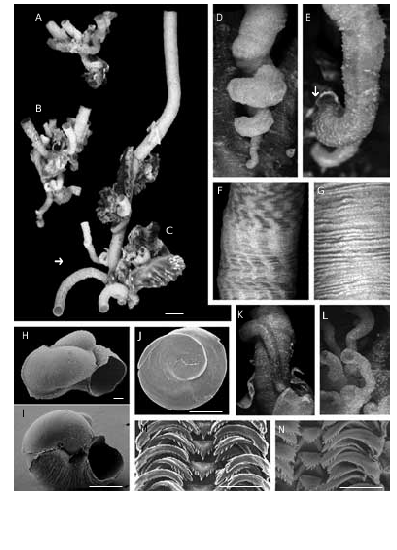
The holotype is part of a bigger cluster of specimens ( Fig. 2C View FIG ). The shell consists of an isolated tube ( Fig. 2C View FIG , arrow), which is cemented and spirally coiled only in its apical portion ( Fig. 2D, E, K, L View FIG ), straight thereafter, as it is typical in this species.
Shell aperture diameters have an average size of 3 to 4 mm. Shells may be completely white or pale yellowish-beige with longitudinal orange-brown lines twisted in some specimens ( Fig. 2F View FIG ). The ornamentation is formed by simple scaly growth stages ( Fig. 2G View FIG ), more evident in juveniles. In some specimens scaly ornamentation organised in long longitudinal ridges, which delimit other completely smooth longitudinal portions. In coloured shells, the scaly ridges correspond to coloured lines. Feeding-tube scars (Keen 1961; Schiaparelli & Cattaneo-Vietti 1999) are practically absent: of more than 250 specimens examined only a juvenile showed a clear arrangement of the tube ( Fig. 2E View FIG ). The shape of the protoconch, with quite disjunct whorls, is uncommon for a vermetid. It has a tip that tends to be erect ( Fig. 2H, I View FIG ) and lacks sculpture; the length is of 600 µm, while the diameter is of 500 µm.
ANATOMY
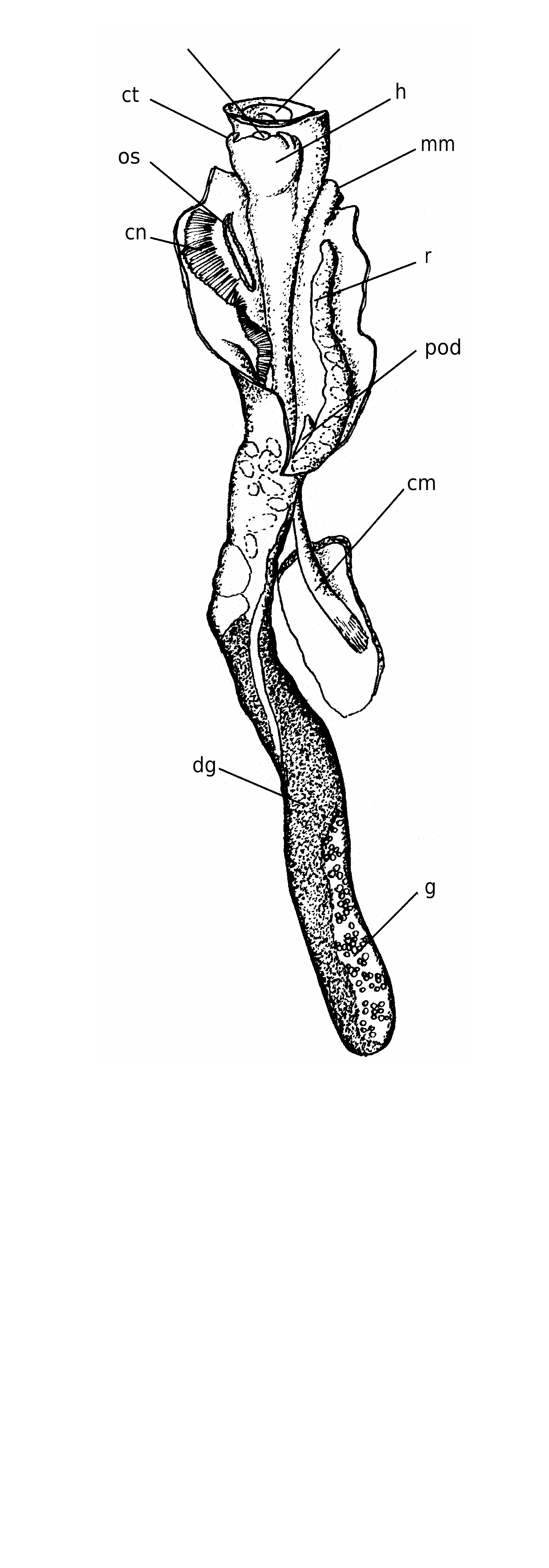
The body ( Fig. 4 View FIG ) is very long and shows no tendency to coil after extraction from the shell. The columellar muscle is well-developed and robust, adapted for deep and fast retreat into the long shell (e.g., Fig. 2C View FIG ).
The foot possesses a slightly concave yellowishbeige operculum that covers the entire surface of the foot. The operculum, which tightly fits the shell aperture (ratio operculum/shell diameter: 8-9/10), has a dirty-white spiral lamina externally ( Figs 2J View FIG ; 3F View FIG ) that contrasts with the darker background of the body colouration. The interior has a low, but distinct attachment scar.
The head is squat with two short cephalic tentacles; the pedal ones small and very difficult to detect, at least in our preserved material. The ctenidium extends for about one-fourth of the total body length and comprises triangular lamellae; at its base an evident osphradium is present. The near absence of pedal tentacles, which are commonly used in vermetids both to spread and to re-collect the mucous net, and the well-developed ctenidium indicate that feeding is exclusively ciliary.
The visceral sac has a digestive acinose portion that is light-brown in colour and shows superficial black spots, both in male and females. The female ovary has racemous blind orange sacs clearly visible due to the transparency of the body in this region; the male gonad is uniformly yellow.
The radula is taenioglossate (2:1:1:1:2) ( Fig. 2M View FIG ). One specimen has an abnormal central cusp in all rows ( Fig. 2N View FIG ).
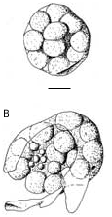
The mantle of the female has no dorsal slit and the egg capsules lie free inside the mantle cavity. In one female we have found two egg capsules containing unsegmented embryos. Only one of the two was not damaged, the other being accidentally squeezed during dissection. The egg capsules are almost spherical and transparent. The diameter of the intact one is of about 600 µm and it is possible to count 21 embryos inside ( Fig. 5A View FIG ). The damaged egg capsule revealed in the inside the presence of smaller globular cells, which are probably polar bodies ( Fig. 5B View FIG ).
The dominant colour of the soft parts beneath the operculum, observed on fresh collected specimens, is a deep orange-red, scattered with yellow small dots on outer mantle margin and around the head ( Fig. 3E View FIG ). The inner mantle margin is yellowish-white, with orange spots that corresponds, on the shell surface, to the coloured scaly processes. A narrow yellow band, regularly interrupted by red squared spots, encircle the foot margin. A yellow “V” encircles the cephalic tentacles.
ECOLOGY
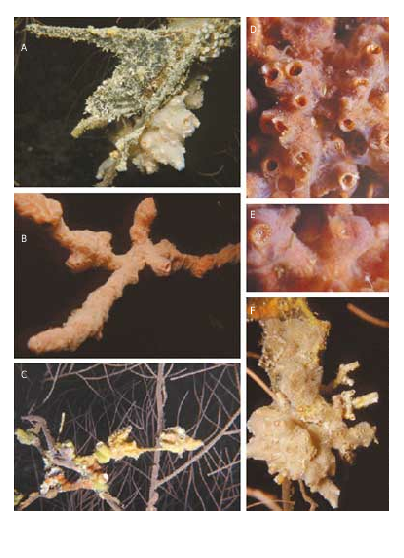
This species forms dense aggregates (> 20 individuals) ( Fig. 3D View FIG ), commonly at 15-20 m depth, on the axial branches of a black coral Anthipates spp. and in association with bivalves as such as Lopha cristagalli , Dendrostrea cf. folium ( Ostreidae ) ( Fig. 3C View FIG ), and Pteria cf. penguin (Röding, 1798) (Pteriidae) ( Fig. 3A View FIG ), which adhere with byssus threads or cement themselves to the branches.
This association of vermetids and bivalves is covered by several species of sponges ( Fig. 3B, D, E View FIG ) and undetermined colonial Ascidiacea ( Fig. 3A, F View FIG ).
Many long (up to 40 cm; diam. 1 cm) feedingtubes characterize the shell of the oldest specimens ( Fig. 3B View FIG ). This length is probably reached in order to avoid competition for food created by the dense branches of the black coral.
Strangely this vermetid does not show any evidence of old abandoned feeding-tubes (Keen 1961) and its growth (except for the very first coils) is practically straight.
Feeding tube scars were erroneously considered as a character of taxonomic value by Keen (1961), as discussed by Bieler (1995), but they are expressed only in some situations (Schiaparelli 1996; Schiaparelli & Cattaneo-Vietti 1999). Their absence could be due to encrusting sponges, which may prevent the mollusc from cutting away the shell edges.
Aggregates of shells up to 1 metre in length have been observed in the Philippines (Springsteen & Leobrera 1986), and their ecology seems to be the same as the Maldivian specimens, as exemplified by the colony figured by Springsteen & Leobrera (1986: pl. 8, fig. 10) growing on a coral or gorgonacean branch.
s o
REMARKS
V. enderi n. sp. is readily distinguishable from the known Indo-pacific vermetid species both for its shell features and the particular ecological niche.
In general, it differs from Hong Kong Vermetus (Vermetus) sp. (Hughes, 1985) in having a bigger operculum and no evident pedal tenctacles; from the western Australian Vermetus (V.) sp. (Hughes
A
1993) in having females without mantle slit and brooding free eggs capsules; from the Hawaiian Vermetus alii Hadfield & Kay, 1972 in having an operculum with only one to two volutions, instead of more than eight, and a protoconch with erected tip.
The South Californian species V. (Thylaeodus) contortus (Carpenter, 1857) (Keen 1961; Morton 1965) and the Mediterranean V. (T.) rugulosus Monterosato, 1878 (Bieler 1995) , both belonging to the same subgenus of this new species, show instead similarities either in anatomical organisation (e.g., brood protection in mantle cavity) and general shape. However they differ in whorls development, sculpture and dimensions.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |