
Symphurus microrhynchus ( Weber, 1913 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5039.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:FB5EDBC1-B2DB-4091-ACDF-C3858F6FCD62 |
|
persistent identifier |
https://treatment.plazi.org/id/094B87CC-FFCF-5E04-B591-3B8CFC2E581B |
|
treatment provided by |
Plazi |
|
scientific name |
Symphurus microrhynchus ( Weber, 1913 ) |
| status |
|
Symphurus microrhynchus ( Weber, 1913) View in CoL
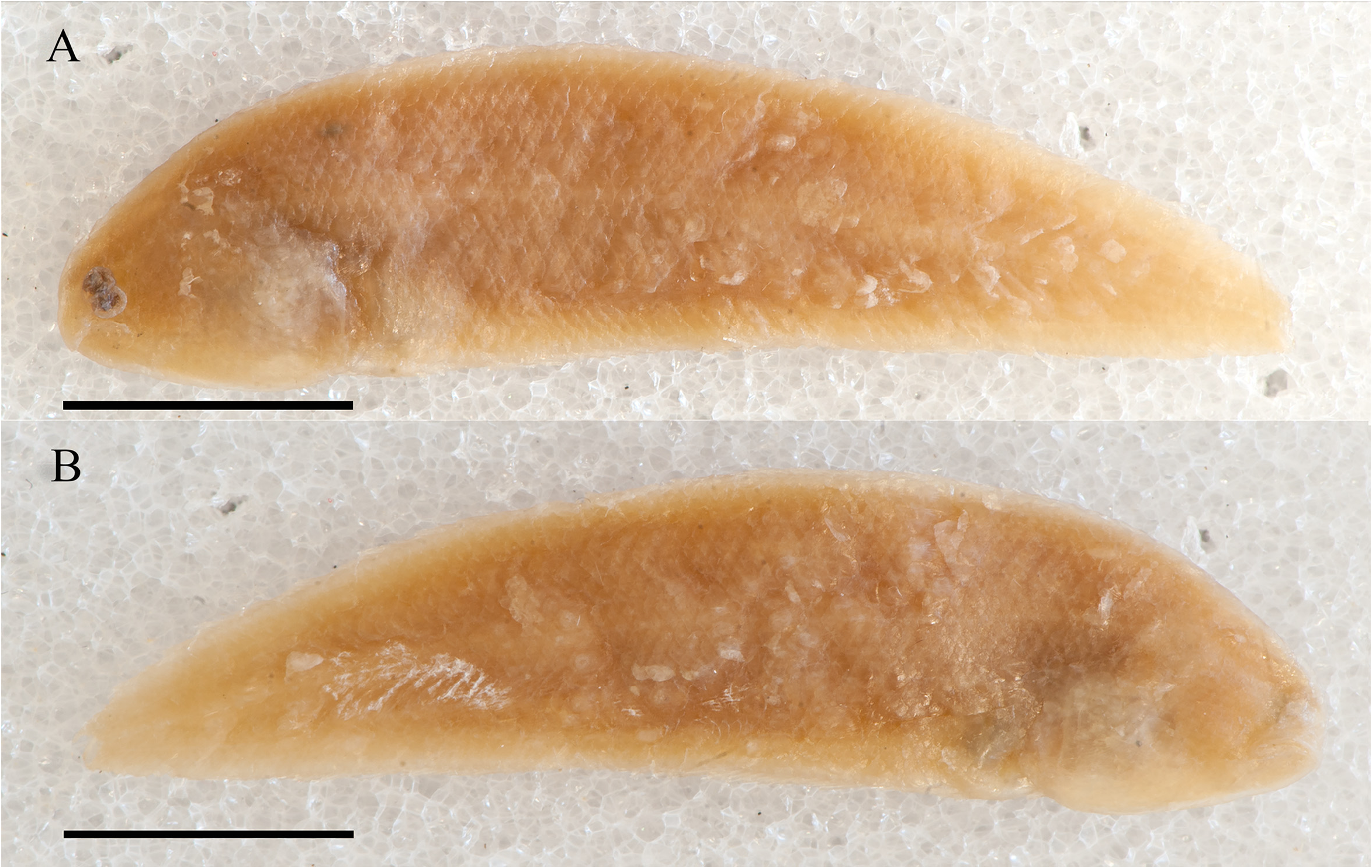
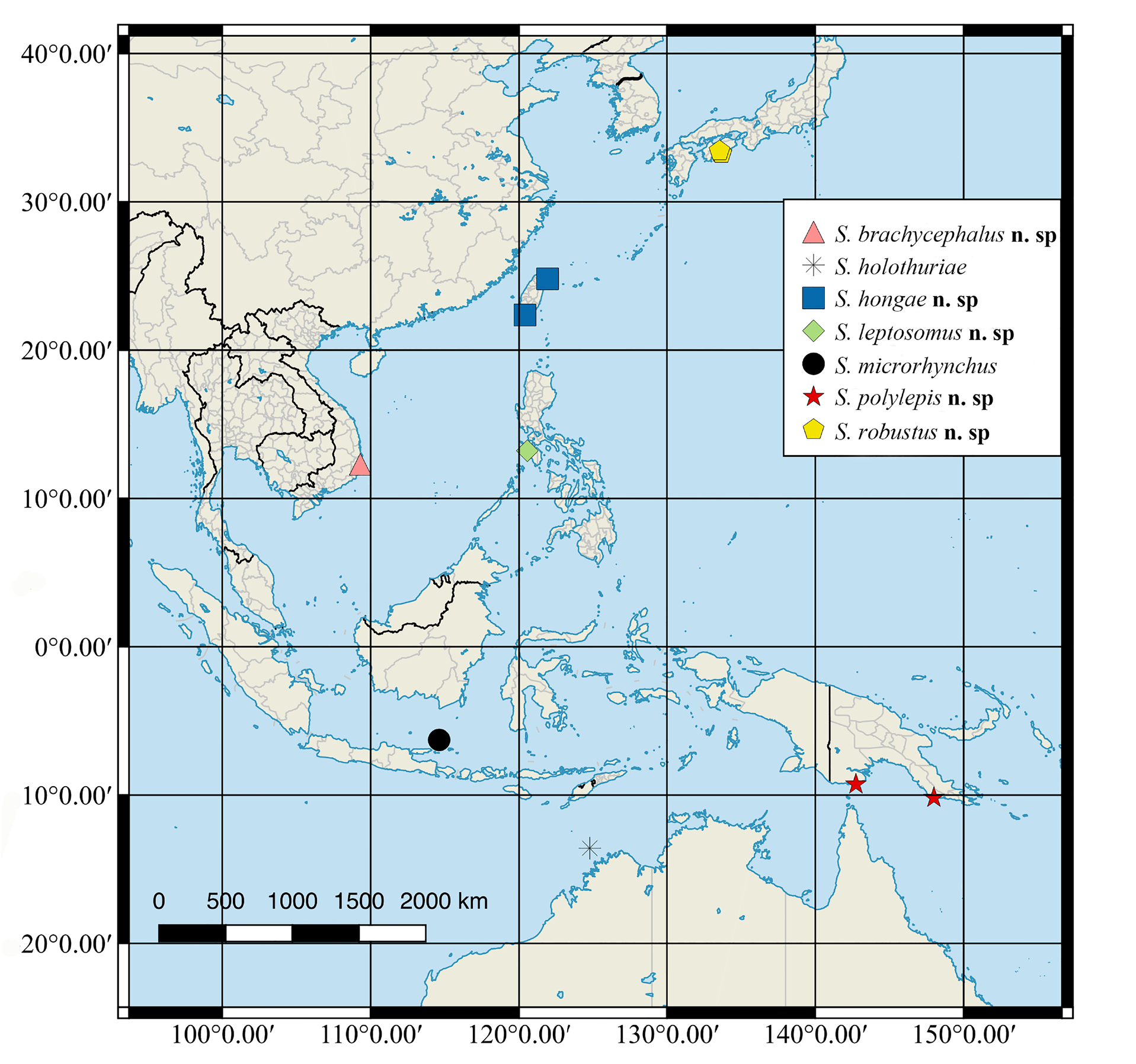
( Figs. 2–3 View FIGURE 2 View FIGURE 3 ; Tables 1–6)
Aphoristia microrhynchus Weber, 1913: 444 View in CoL , Fig. 85 (Java Sea GoogleMaps , 6°16.5’S, 114°37’E, Siboga View in CoL station 319, depth 82 m, Holotype: ZMA 108193 View Materials ).
Symphurus microrhynchus View in CoL . Weber and de Beaufort, 1929: 209, 211 (redescription; transfer to Symphurus View in CoL ; in identification key with S. regani View in CoL and S. gilesii View in CoL ). Chabanaud 1939: 27 (listed). Koumans 1953: 214 (one specimen from Indonesia, without descriptive information). Chabanaud 1955b: 45 (redescription; comparisons with S. holothuriae View in CoL and S. trifasciatus View in CoL ; S. holothuriae View in CoL a possible synonym). Munroe 1992: 361 (provided informative counts and compared with congeners). (?) Dufour et al. 1996: 415 (several specimens metamorphosing to juvenile stage; mentioned in Munroe & Marsh 1997 as possibly this species). Munroe & Marsh, 1997: 191 (in part) (redescription based on specimens of several different species; compared with S. holothuriae View in CoL and S. trifasciatus View in CoL ). Lee et al. 2014: 564 (in part) ( S. microrhynchus sensu Munroe & Marsh 1997 View in CoL composite mix of several different species; comparisons of S. microrhynchus sensu Munroe & Marsh 1997 View in CoL with S. leucochilus View in CoL ). Lee et al. 2017: 72 (in part) ( S. microrhynchus sensu Munroe & Marsh 1997 View in CoL composite mix of several different species; comparisons of S. microrhynchus sensu Munroe & Marsh 1997 View in CoL with S. longirostris ).
Symphurus microrhynchus View in CoL (not Weber). Voronina et al. 2016: 401 (21 specimens, Vietnam; meristics, color pattern; diagnostic features; possibility of a cryptic species; distribution) .
Holotype: ZMA 108193 View Materials , 42.2 mm SL, immature female, Indonesia: Java Sea , 6°16.5’S, 114°37’E, Siboga station 319, depth 82 m, 22 Feb 1900. GoogleMaps
Diagnosis. Symphurus microrhynchus is distinguished from all congeners by the combination of: a 1–2–2–2–2 ID pattern; 12 caudal-fin rays; 9 (3 + 6) abdominal vertebrae; 48 total vertebrae; 4 hypurals; 87 dorsal-fin rays; 73 anal-fin rays; 64 longitudinal scale rows; 25 transverse scale rows; and 15 scale rows on the head posterior to the lower orbit; body elongate (BD= 24.9% of SL); preanal length long (PAL= 28.1% of SL); upper head lobe much smaller than lower head lobe; head long (HL= 24.2% of SL), longer than head width (HW/HL= 0.97); postorbital length long (POL= 76.5% of HL); snout short (SNL= 10.4% of HL, SNL/ED= 1.25), slightly rounded anteriorly; dorsal-fin origin located at vertical posterior to upper eye; predorsal length short (PDL= 18.1% of HL); eye relatively small (ED= 8.4% of HL); eyes unequal in position with anterior margin of upper eye greatly in advance of anterior margin of lower eye; fleshy ridge well developed on posterior part of ocular-side lower jaw; with membrane covering upper and lower eyes; with continuous, fleshy membrane connecting anterior nostril with lower part of eye; ocular-side pigmentation uniformly yellowish-brown; blind-side pigmentation uniformly yellow; both sides with obvious dermal spots on bases of anteriormost dorsal and anal fins.
Description. Symphurus microrhynchus is likely a dwarf species. This species is known from two specimens. The only specimen available to this study was the holotype, an immature female measuring 42.2 mm SL ( Fig. 2 View FIGURE 2 ). Meristic characters are summarized in Tables 2–5. ID pattern 1–2–2–2–2. Caudal-fin rays 12. Dorsal-fin rays 87. Anal-fin rays 73. Pelvic-fin rays 4. Total vertebrae 48; abdominal vertebrae 9 (3 + 6). Hypurals 4. Longitudinal scale rows 64. Scale rows on head posterior to lower orbit 15. Transverse scale rows 25.
Proportions of morphometric features presented in Table 6. Body elongate; maximum depth in middle of body, approximately at point of insertion of first anal-fin ray, body with gradual taper posterior to midpoint. Preanal length much longer than body depth. Head moderately long and narrow; head width shorter than head length (HW/HL= 0.97). Upper head lobe much smaller than lower head lobe (UHL/LHL= 0.78), and much shorter than postorbital length. Lower lobe of ocular-side opercle wider than upper opercular lobe; posterior margin of lower lobe projecting slightly beyond posterior margin of upper opercular lobe. Snout short, slightly rounded anteriorly, its length slightly greater than eye diameter (SNL/ED= 1.25). Dermal papillae present and well developed on blind side of snout and chin. Ocular-side anterior nostril tubular and short, when depressed posteriorly usually not reaching anterior margin of lower eye. Ocular-side posterior nostril a small, rounded tube located on snout just anterior to interorbital space. Blind-side anterior nostril tubular and relatively short, easily distinguishable from dermal papillae; blind-side posterior nostril a shorter and wider, posteriorly directed tube situated posterior to vertical at posterior margin of jaws. Jaws long and slightly arched; upper jaw length slightly longer than snout length; posterior margin of upper jaw extending to vertical through posterior margin of lower eye. Ocular-side lower jaw with well-developed fleshy ridge on posterior portion. Cheek depth relatively wide, slightly longer than snout length. Eyes small, oval, unequal in position, upper eye greatly in advance of lower eye. Pupillary operculum absent. Dorsal-fin origin located at vertical posterior to upper eye; predorsal length relatively short. Anteriormost dorsal-fin rays obviously shorter than more posterior fin rays. Scales absent on both sides of dorsal- and anal-fin rays. Pelvic fin long; longest pelvic-fin ray, when extended posteriorly, usually reaching base of sixth anal-fin ray. Posteriormost pelvic-fin ray connected to anal fin by delicate membrane. Caudal fin broken, unsure if base of fin with rows of ctenoid scales present or not. Scales on both sides of body numerous, strongly ctenoid.
Teeth present and recurved slightly inwards on all jaws, but better developed on blind-side jaws. Ocular-side premaxilla and dentary with single row of sharply pointed, well-developed teeth. Blind-side premaxilla with two rows of sharp, recurved teeth. Blind-side lower jaw with three rows of well-developed teeth.
Pigmentation. Pigmentation of live or freshly collected S. microrhynchus unknown. From preserved holotype ( Fig. 2 View FIGURE 2 ), ocular-side background pigmentation generally yellowish-brown, without pepper-dot melanophores in dermis overlying pterygiophores. External surface of abdominal area dark brown. Background pigmentation of ocular side of head generally similar to that on body. Ocular side of snout light yellow. Dermal spots present at bases of anteriormost dorsal-fin rays and also on isthmus. Outer surface of ocular-side opercle yellowish-brown, margin of opercle yellowish-brown, with same background pigmentation as that on body and head. Inner surface of ocularside opercle and isthmus with small light-brown dots, ocular-side lips and chin region uniformly yellow. Ocularside anterior nostril light yellow. Upper aspects of eyes and eye sockets bluish-black; pupils bluish-black, lighter than pigmentation of eye sockets. Blind side uniformly yellow. No median longitudinal row of darkly pigmented melanophores evident along vertebral axis. Outer surface of blind-side opercle light yellow similar to background pigmentation. Inner surface of blind-side opercle unpigmented. Fin rays of dorsal, anal, and pelvic fins uniformly yellowish-brown; basal regions of fin rays and membranes covering fin rays light yellow.
Distribution. The holotype of S. microrhynchus was collected at a depth of 82 m in the Java Sea, off Indonesia ( Fig. 3 View FIGURE 3 ). Koumans (1953) reported a second specimen (RMNH 20305, 27.0 mm SL) identified as this species, from the Savu Sea near Koepang, Indonesia. Munroe & Marsh (1997) examined this specimen and agreed with Kouman’s identification. Reports of S. microrhynchus from locations other than Indonesia that appear in Munroe & Marsh (1997), including Zanzibar, Tanzania; Papua New Guinea (see also Kailola 1971); French Polynesia; northwest Australia; and the Arafuru Sea, as well as specimens from Vietnam ( Voronina et al. 2016) are not S. microrhynchus , but are based on other species (see species accounts below).
Remarks. Symphurus microrhynchus was described from a single specimen collected in 1900 off Indonesia ( Weber 1913). Weber compared and distinguished his new species from two deepwater western Indian Ocean species, Aphoristia trifasciatus Alcock and A. septemstriatus Alcock (both species now placed in Symphurus ), noting especially the fewer numbers of longitudinal scales in his new species (approximately 68) compared with the much higher values in these others. Weber also mentioned some pigmentation features of the preserved holotype of his new species including the statement that the specimen was apparently faded, but that the ocular side was darker than the blind side and had darker, round, scattered spots along bases of the dorsal and anal fins. Chabanaud (1955b), in his redescription of this species, also noted the specimen was devoid of pigment except for these melanophores. Weber commented that the holotype also had weak indication of about five transverse bands. Evidence of these interrupted bands and also some large blotches appear in the figure accompanying the original description of this species. Weber described the blind side of the holotype as light brown, but did not mention any pepper-dot melanophores overlying pterygiophore regions of blind sides of the dorsal and anal fins, nor did he describe any large melanophores along the vertebral axis on the blind side.
Munroe & Marsh (1997) redescribed S. microrhynchus based on a composite series of specimens collected from geographic locations including Zanzibar, Tanzania, Vietnam, New Guinea, Indonesia, French Polynesia, and Australia. They considered S. microrhynchus to be a widespread species, but new data (see below) reveals that at least five other species were included in their account for S. microrhynchus . Lee et al. (2014; 2017) also confirmed that more than a single species was present among material identified as S. microrhynchus in Munroe & Marsh (1997). In fact, among specimens examined by Munroe & Marsh (1997), approximately six nominal species (including S. holothuriae ) were included.
Re-examination of the holotypes of S. microrhynchus and S. holothuriae , led Lee et al. (2014; 2017) to discover useful characters to distinguish these two nominal species, and these authors suggested that these two nominal species should both be recognized as valid species based on differences in several characters, including those in ID pattern and counts for total vertebrae. These and other differences between S. microrhynchus and S. holothuriae are provided in the Comparisons section below.
Another finding provided by Lee et al. (2014; 2017) is discovery that S. microrhynchus and S. holothuriae have much lower scale counts than those of other specimens identified as S. microrhynchus by Munroe & Marsh (1997). This finding identified two groups of species within the S. microrhynchus species complex: a high scale count group and a low scale count group.
Re-examination of the type specimens of S. microrhynchus and S. holothuriae by M.Y. Lee for the present study also revealed some incongruous information concerning morphometric values between Lee’s unpublished data and that of Munroe & Marsh (1997). In their study, Munroe & Marsh (1997) recorded the UHL as being equal to the LHL in S. microrhynchus , in contrast, the LHL was reported to be larger than the UHL in S. holothuriae . Re-examination of the type specimens by Lee during his work conducted at the USNM actually revealed that these data are reversed. This fact was verified during several re-examinations of the types and also from data available from photographs. The incongruity between measurements made in the re-examination and that reported in Munroe & Marsh (1997) only happened for the UHL & LHL measurements, and it appears that these differences resulted from interchanging data reported for these two species. We suggest the revised data of M.Y. Lee are more accurate than that reported in Munroe & Marsh (1997), and thus these data are utilized in the redescriptions of S. microrhynchus above and for that of S. holothuriae , which follows below.
Symphurus microrhynchus is only known from the preserved holotype and one other preserved specimen (Koumans 1951). Additional specimens are needed to gain robust data, including both morphological and molecular data, for distinguishing this species. Not only would newly collected specimens provide additional information for understanding intraspecific variation in morphological features, but these specimens would also provide the source for genetic information, so necessary for integrating morphological and molecular approaches for resolving taxonomic issues and discovering systematic relationships of S. microrhynchus and morphologically similar species. Additional specimens will also provide better understanding of the ecology, abundance, demographics, and life history of this species.
Comparisons. Symphurus microrhynchus is one of three species in the S. microrhynchus species complex featuring lower scale counts. Other members included in this species group are S. holothuriae and S. longirostris . Comparisons of S. microrhynchus and S. holothuriae are limited because each of these nominal species is known only from their holotypes.
Symphurus microrhynchus differs from S. holothuriae in having a 1–2–2–2–2 ID pattern (vs. 1–2–3–2–2 pattern in S. holothuriae ). Whether this difference in ID pattern is a real distinction between these two nominal species, or merely represents intraspecific variation within S. holothuriae (see comments below), will require additional specimens of both species to determine their predominant ID patterns. The holotype of S. microrhynchus has 48 total vertebrae compared with 46 in the holotype of S. holothuriae . Values for this character may also be a species level diagnostic feature between these two species, but again more specimens of each species are needed to determine intraspecific variation of vertebral numbers in these species and whether values for this character are useful in diagnosing these two species.
of Indo-Pacific.
Among morphometric features, S. microrhynchus differs from S. holothuriae with respect to the width of the lower and upper head lobes. In S. microrhynchus , the lower head lobe is slightly wider (LHL= 13.1% of SL) compared with that of S. holothuriae (LHL= 11.6% of SL). In contrast, the upper head lobe of S. microrhynchus is slightly narrower (UHL= 10.3% of SL) compared with that of S. holothuriae (UHL= 11.7% of SL). These differences are reflected in the ratios of UHL/LHL in the two species (0.78 in S. microrhynchus vs. 1.01 in S. holothuriae ). In other species of Symphurus observed in previous studies ( Lee et al. 2009b; 2013; 2014; 2017), intraspecific variation in the ratio between the UHL and LHL is usually stable and thus this feature is usually useful to separate species into two groups: those with UHL/LHL ≤1 versus those with UHL/LUL>1. In S. holothuriae , the UHL=11.7% SL is slightly larger than the LHL (11.6% of SL).
Symphurus microrhynchus is similar to S. longirostris , the third member of the low scale count group of the S. microrhynchus species complex, in general body shape, and counts of dorsal-, anal- and caudal-fin rays, total vertebrae, longitudinal scales, and transverse scales ( Table 6). Symphurus microrhynchus differs from this species in having more rows of scales on the head posterior to the lower eye (15 vs. 11–14 in S. longirostris ). These species differ with respect to several morphometric features ( Table 6), including that S. microrhynchus has a longer and wider head (vs. shorter and narrower in S. longirostris ); a shorter SNL (vs. longer); smaller ratio of SNL/ED (larger ratio in S. longirostris ); shorter lower jaw (vs. longer); relatively longer POL; more anterior insertion of the 1 st dorsal-fin ray in S. microrhynchus , which results in a shorter PDL (18.1% of HL) in this species compared with that of S. longirostris (PDL= 23.8–32.7% of HL); and the lower head lobe of S. microrhynchus is larger than the upper head lobe (UHL/LHL= 0.78), whereas in S. longirostris the lower head lobe is smaller than the upper head lobe (UHL/LHL= 1.01–1.20). Other differences between S. microrhynchus [sensu this study] and S. longirostris were discussed in Lee et al. (2017).
Symphurus microrhynchus differs from the five species of the S. microrhynchus species group featuring high scale counts ( S. brachycephalus n. sp., S. hongae n. sp., S. leptosomus n. sp., S. polylepis n. sp., and S. robustus n. sp.) by its fewer number of rows of longitudinal (64 vs. 72–94) and transverse scales (25 vs. 26–37 in these others). And, in S. microrhynchus , head width is shorter than head length (HW/HL= 0.97), whereas, in the five species with high scale counts, their heads are wider than long with HW/HL ratios ranging from 1.02–1.20 for these species.
Symphurus microrhynchus has the same ID pattern (1–2–2–2–2) and overlapping counts for dorsal-fin rays (87 vs. 86–92), anal-fin rays (73 vs. 73–80), and total vertebrae (48 vs. 48–51) as those found in S. monostigmus and S. leucochilus . However, S. microrhynchus is readily differentiated from both S. monostigmus and S. leucochilus in having a membranous connection between the ocular-side anterior nostril and the lower eye, and this species also has a fleshy ridge on the ocular-side lower jaw; both features are absent in S. monostigmus and S. leucochilus . In S. microrhynchus , head length is nearly equal to its width (HW/HL= 0.97) versus head width greater than head length (HW/HL= 1.07–1.42) in these other two species. Additionally, S. microrhynchus has fewer rows of longitudinal (64 vs. 75–93) and transverse scales (25 vs. 32–38) than is found in these others. Symphurus microrhynchus differs further from S. monostigmus in having 12 (vs. 14) caudal-fin rays and 4 (vs. 5) hypurals, and in lacking the pigment spot on the abdomen that features so prominently in S. monostigmus .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Symphurus microrhynchus ( Weber, 1913 )
| Lee, Mao-Ying & Munroe, Thomas A. 2021 |
Symphurus microrhynchus
| Voronina, E. P. & Prokofiev, A. M. & Prirodina, V. P. 2016: 401 |
Symphurus microrhynchus
| Lee, M. - Y. & Kai, Y. 2017: 72 |
| Lee, M. - Y. & Munroe, T. A. & Shao, K. - T. 2014: 564 |
| Munroe, T. A. & Marsh, B. N. 1997: 191 |
| Dufour, V. & Riclet, R. & Lo-Yat, A. 1996: 415 |
| Munroe, T. A. 1992: 361 |
| Chabanaud, P. 1955: 45 |
| Koumans, E. R. 1953: 214 |
| Chabanaud, P. 1939: 27 |
| Weber, M. & de Beaufort, L. F. 1929: 209 |
Aphoristia microrhynchus
| Weber, M. 1913: 444 |