
Taito adrik, Friedrich & Lehmann, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4729.1.7 |
|
publication LSID |
lsid:zoobank.org:pub:2356D7B2-C8AB-4AAC-B0B3-714FFB6621F5 |
|
persistent identifier |
https://treatment.plazi.org/id/F177DAEC-D323-46ED-9DBE-B0C6794775DA |
|
taxon LSID |
lsid:zoobank.org:act:F177DAEC-D323-46ED-9DBE-B0C6794775DA |
|
treatment provided by |
Plazi |
|
scientific name |
Taito adrik |
| status |
sp. nov. |
Taito adrik View in CoL sp. nov.
( Figs. 1–7 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
urn:lsid:zoobank.org:act:F177DAEC-D323-46ED-9DBE-B0C6794775DA
Etymology. The specific name is an indeclinable noun in apposition, given in honour of the outstanding arachnologist Adriano B. Kury (nickname “Adrik”), who established the genus Taito , together with Carla M.L. Barros, in 2014.As professor and senior curator of arachnids in the Museu Nacional/UFRJ, Rio de Janeiro, Brazil, he lost most of his invaluable collections and fruits of his many field trips through the fire catastrophe in September 2018.
Type material. PERÚ, Huánuco Department, Puerto Inca Province, Yuyapichis District , Rio Yuyapichis , ACP Panguana (9°37’S, 74°56’W, 230 m a.s.l.): holotype male ( MUSM 0513700 ), 21.iv.–04.v.2018 (E. Diller) GoogleMaps ; 1 female paratype ( ZSMA20190295 ), 02.–18.x.2009 (E.-G. Burmeister) ; 3 male and 1 female paratypes ( MUSM 0513706 /707, ZSMA20190294 , ZMH-A0002240 ), 20.ix.–07.x.2013 (S. Friedrich & F. Wachtel) ; 2 female paratypes ( ZSMA20190289 /290), 01.–21.v.2015 (S. Friedrich, F.Wachtel & M. Steinherr) ; 2 male paratypes ( ZMH-A0002241 , MHNG-ARTO-18641), 23.iv.–09.v.2016 (S. Friedrich, F. Wachtel & D. Hauth) ; 1 male paratype ( ZSMA20190296 ), 22.ix.–10.x.2017 (S. Friedrich, F. Wachtel, D. Hauth & T. Lehmann) ; 1 female paratype (MHNG-ARTO-18642), 22.ix.–10.x.2017 (E.-G. Burmeister); 2 male and 3 female paratypes ( MNRJ 6033–036 View Materials , ZSMA20190297 ), 21.iv.– 04.v.2018 (E. Diller) ; 7 male and 11 female paratypes ( ZSMA20190291–293 /298–304, MUSM 0513701–705 /708– 710), 24.viii.–06.ix.2018 (E. Diller). All specimens were collected at night on the ground in primary evergreen lowland rainforest .
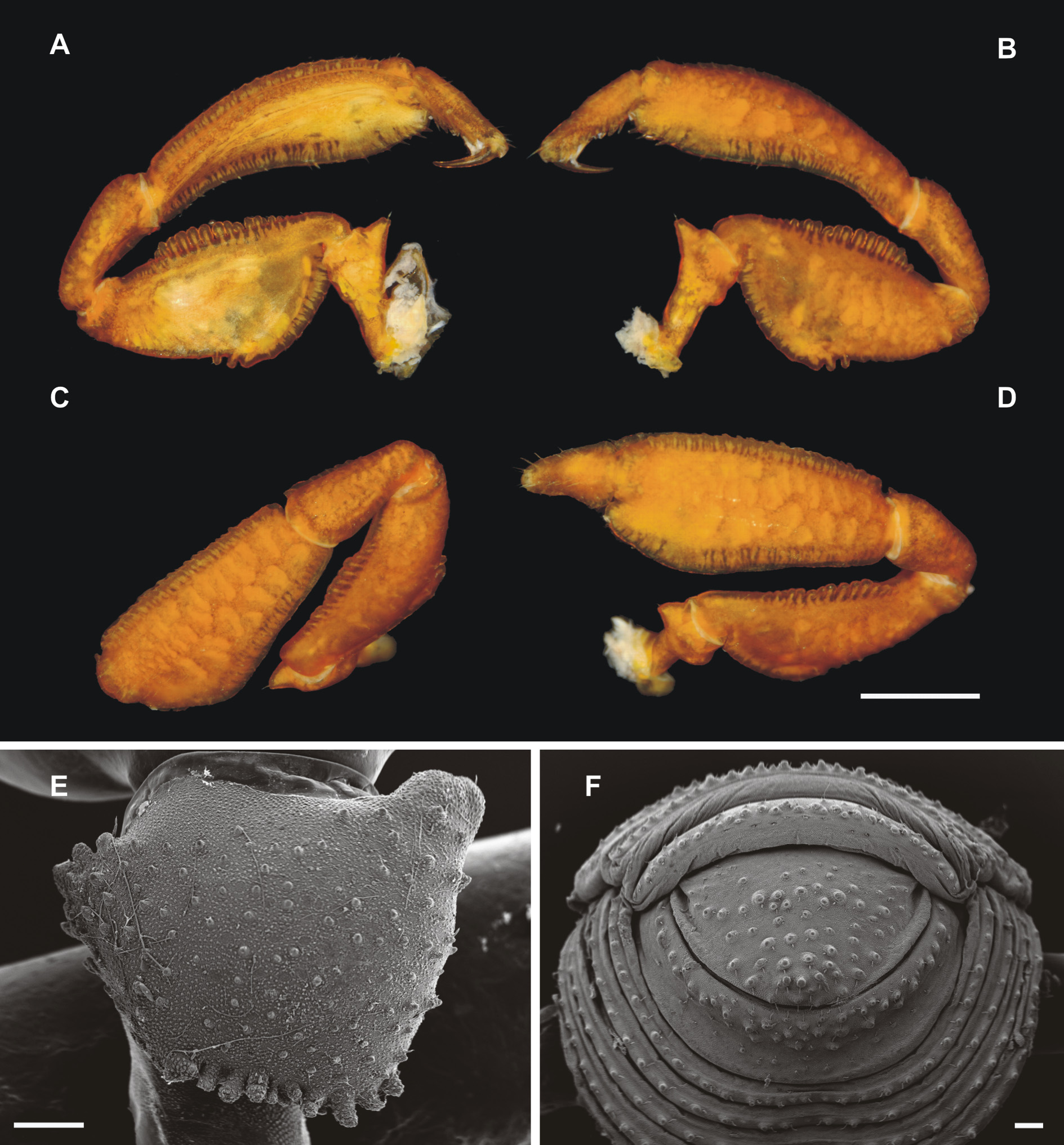
Diagnosis. Dorsal scutum sturdy (average ratio length/width ca. 1.18 in females, ca. 1.21 in males), β-type (normal β) ( Kury et al. 2007) and not βL-type (elongate β) ( Kury & Medrano 2016) ( Fig. 2 A View FIGURE 2 ). Equuleus butterflyshaped with discrete feet and arms, and elongated, slim horns ( Figs. 1 A View FIGURE 1 ; 2 A, C View FIGURE 2 ; 3 View FIGURE 3 ); two whitish blots on minute tubercles at posterior margin of scutal area III (in some specimens two smaller additional blots beside) ( Figs. 1 A View FIGURE 1 ; 2 A, C View FIGURE 2 ). Groin warts ( Kury & Barros 2014) on dorso-proximal part of coxa IV ( Fig. 2 A, B View FIGURE 2 ), coxa IV with dorsal triangular apophysis ( Fig. 2 F View FIGURE 2 ). Leg IV of male: femur IV gently curved with comb of five to six curved apophyses on distal-prolateral side; patella IV with coarse granules and one single apophysis proximal-retrolateral; tibia IV unarmed, covered with granules ( Figs. 2 E, F View FIGURE 2 ; 5 View FIGURE 5 ).
Distribution. Only known from the type locality ( Fig. 1 B View FIGURE 1 ). This extends the distribution range of the genus Taito to the south-west. So far, the genus was known from the Upper Amazon Basin up the eastern slope of the Andes. Now, the distribution reaches to a territory west of the Ucayali River, into the Pachitea Basin, which is located between the eastern slopes of the Andes and the Sira Mountains ( Fig. 1 B View FIGURE 1 ).
Description. Male holotype (MUSM 0513700)
Color (in ethanol) ( Fig. 2 View FIGURE 2 ). Body medium brown, densely covered with small, lighter orange-brown islands. Appendages also showing this pattern, but a little bit lighter. Equuleus on dorsal scutum and interrupted stripe on coda nearly white. Shape of equuleus very consistent ( Fig.3 View FIGURE 3 ).
Measurements. CL = 2.3, CW = 3.6, AL = 4.2, AW = 5.7; legs I to IV: Table 2 View TABLE 2 .
Dorsum ( Figs. 2 A, D View FIGURE 2 ; 4 F View FIGURE 4 ). Dorsal scutum sturdy (ratio ca. 1.19), maximum width at scutal area III, thickened laterals with granulate surface; two minute tubercles on area III. Posterior margin of scutum sub-straight in dorsal view. Posterior margin of free tergite I also sub-straight, that of free tergite II weekly convex, and of free tergite III strongly convex ( Fig. 2 A View FIGURE 2 ). All free tergites with a row of transverse granules. Anal operculum unarmed, with two dorso-lateral indentations, covered with granules ( Figs. 2 D View FIGURE 2 ; 4 F View FIGURE 4 ).
Venter ( Figs. 2 B View FIGURE 2 ; 4 F View FIGURE 4 ). Coxae I–II nearly transverse, parallel and sub-equal in size. Coxa III larger. Coxa IV much stronger and slanted backwards. Anterior part of genital operculum elliptical, posterior margin truncated. Stigmatic area y-shaped with large elliptical stigmata. Stigmatic area deeply concave relative to the greatly bulging coxa IV. Free sternites I–VI strongly concave, with a dense row of granules.
Chelicerae ( Fig. 4 E View FIGURE 4 ). Basichelicerite with an ectal row of six tubercles; three anterior tubercles large and connected, and a posterior row of 12 granulate tubercles with nearly equal size. One single large and rounded anteromesal process.
Pedipalps ( Fig. 4 View FIGURE 4 A–D). Trochanter with a single ventral spine. Femur with dorsal proximal keel with two anterior teeth, ventral row of 17 teeth, the middle one largest. Tibia with an apophysis at distal end, near tarsus.
Legs ( Figs. 2 E, F View FIGURE 2 ; 5 View FIGURE 5 ). Legs I–III granulous and unarmed. Femur IV gently curved both laterally and dorsoventrally, finely granulous; it is armed with a prolateral comb of six posteriorly curved spine-shaped apophyses, at distal end. Patella IV covered with coarse granulation and armed with a single retrolateral apophysis at the proximal end. Tibia IV unarmed with granulous surface. Tarsal count: 6–6/14–14/7–7/9–9. Measurements of leg segments provided in Table 2 View TABLE 2 .
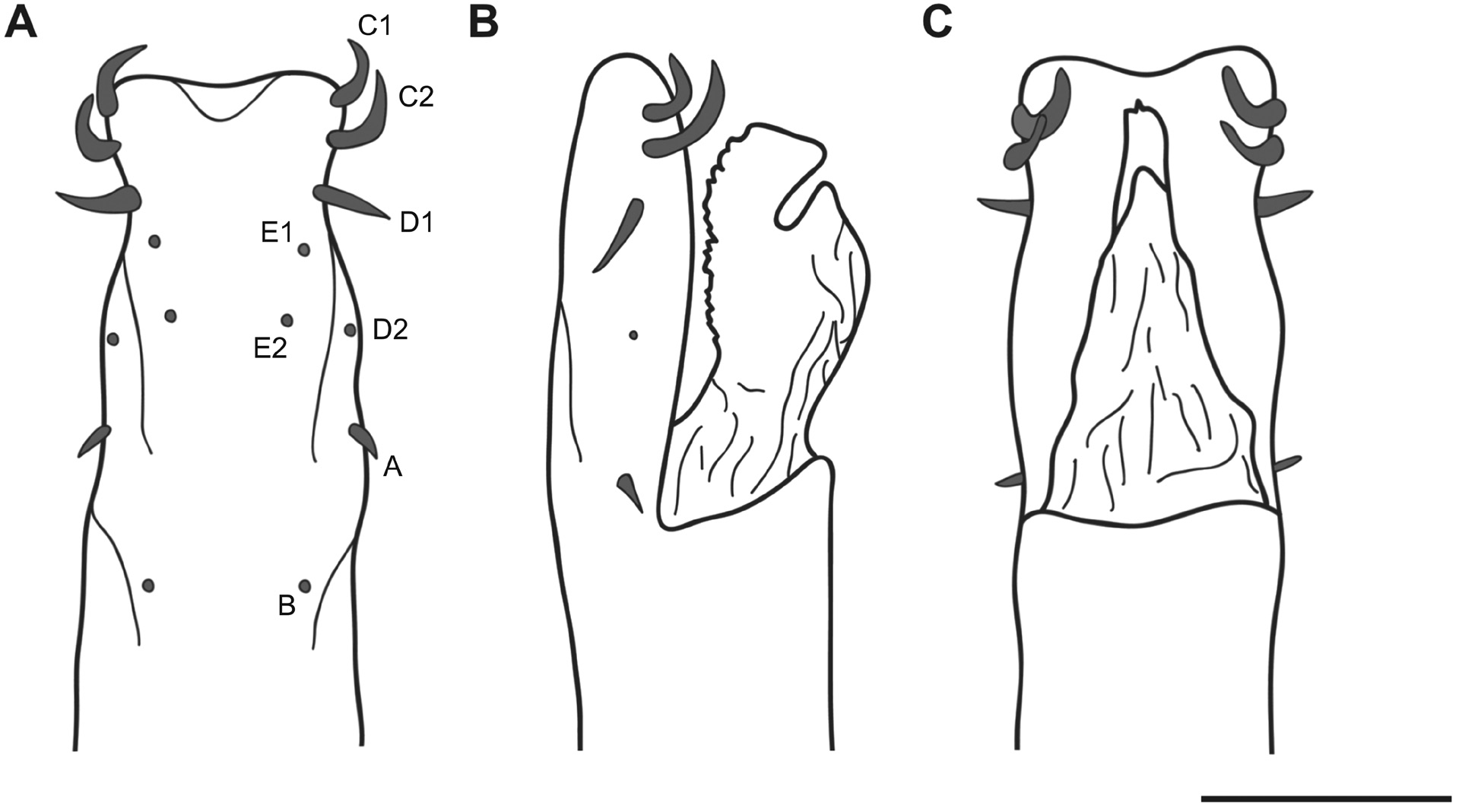
Male genitalia ( Figs. 6 View FIGURE 6 A–C; 7 A–H). Penis: slender trunk with distal bulge. Dorsal process of glans thumb-like and smooth. Glans sac with cellulite texture. Stylus covered with spiny tubercles on its ventral border. Ventral plate with macrosetae C1 and C2 large, curved and spatulate. D1 short and straight, or large and curved in a paratype (see Discussion). D2 reduced. Only one pair of macrosetae A and B. A small and straight, B reduced, E1 and E2 reduced. Microsetae (ms) on ventral plate with a wide central gap and made up of type 4 ms distally, which gradually become type 1 ms proximally.
Comparisons. In several Taito species, femur IV is armed with a row/comb of a few distal spines; in T. medinae , there are eight spines, while in T. spaceinvaders , T. osmari , T. insperatus , and T. oblongatus there are seven spines. Only in T. adrik sp. nov. the number of distal spines is six ( Fig. 2 E, F View FIGURE 2 ; 5 View FIGURE 5 ). Generally, the armature of leg IV in T. adrik sp. nov. is similar to two closely related species, T. insperatus and T. oblongatus . However, these species differ in having dorsal scutum elongated, while it is sturdy in T. adrik sp. nov., a feature only shared with T. serriperna and T. kawaiikei . These species in turn are easily separated from T. adrik sp. nov. especially by the armature of leg IV (apophyses on coxa, femur, and patella). With respect to coxa IV, in T. adrik sp. nov. it has a single small apophysis ( Fig. 2 A, F View FIGURE 2 ), just like in T. insperatus , T. kakera , T. medinae , T. oblongatus , T. osmari , T. rorschachi , and T. spaceinvaders ; in contrast, T. galaga and T. honda have a single large apophysis, and T. juruensis , T. kawaiikei , and T. serriperna have a double apophysis. Patella IV is unarmed in most species; in a few species, this article is covered with acuminate/setiferous tubercles ( T. galaga , T. kakera , T. osmari , and T. rorschachi ). T. adrik sp. nov. is the only species with a single retrolateral conspicuous apophysis ( Fig. 2 E, F View FIGURE 2 ; 5 View FIGURE 5 ). T. galaga and T. rorschachi are the only species that share a butterfly-shaped equuleus with the new species. In T. adrik sp. nov. the design of the butterfly is most similar to T. galaga , but only in the new species the equuleus has elongated, slim horns reaching the ocularium ( Fig. 1 A View FIGURE 1 ; 2 A, C View FIGURE 2 ; 3 View FIGURE 3 ). Moreover, T. adrik sp. nov. differs from T. galaga and T. rorschachi in several other features (scutal area III, anal operculum, and armature of leg IV). In the genus Taito , scutal area III has either high spines ( T. galaga and T. honda ) or small granules/minute tubercles (all other species, including T. adrik sp. nov., Fig. 2 A, C, D View FIGURE 2 ). Finally, the dorsal anal operculum is either unarmed ( T. adrik sp. nov., Fig. 2 D View FIGURE 2 ; 4 F View FIGURE 4 ), as well as T. kakera , T. honda , T. insperatus , T. oblongatus , T. kawaiikei , T. juruensis , and T. serriperna ), or armed with a median spine, a blunt protuberance or a crown of tubercles ( T. spaceinvaders , T. galaga , T. honda , T. medinae , T. osmari , T. rorschachi , and T. spaceinvaders ); however, only in T. adrik sp. nov. the anal operculum has two dorso-lateral indentations, which can be seen with the SEM and were not described in all other Taito species so far.
The new species can be inserted in the key of Kury & Barros (2014:20), at the dichotomy 7(2) (then adjusting the numbering of dichotomies by changing the former 7(2) to 8(7), and so on), as follows:
7(2) Equuleus butterfly-shaped with elongated, slim horns ( Fig. 2 A View FIGURE 2 , 3 View FIGURE 3 aa–bs); femur IV armed with a comb of 6 distal spines ( Fig 2 E, F View FIGURE 2 ; 5 View FIGURE 5 ); patella IV armed with a single retrolateral conspicuous apophysis ( Fig 2 E, F View FIGURE 2 ; 5 View FIGURE 5 )............ Taito adrik sp. nov. Equuleus H- or easel-shaped; femur IV with a comb of 7 or 8 distal spines; patella IV unarmed....................... 8
COI barcoding. Specimen data and DNA sequences of the studied species are available from BOLD ( Ratnasingham & Hebert 2007). The genetic divergences between the six sequenced specimens (holotype and five paratypes) range between 0.0%, and 2.0% (average 0.8%, see Table 1 View TABLE 1 ). A search in BOLD revealed specimens of the cosmetids Cynorta sp. (86.49% similarity), Vonones sp. (85.71% similarity), and Paravonones sp. (85.65% similarity) as the closest matches.





| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |