
Tatuidris tatusia Brown and Kempf 1968
|
publication ID |
https://doi.org/ 10.5281/zenodo.282479 |
|
publication LSID |
lsid:zoobank.org:pub:F7AF89A9-D1B5-4FFF-BD8A-D31BF24EB725 |
|
DOI |
https://doi.org/10.5281/zenodo.5631214 |
|
persistent identifier |
https://treatment.plazi.org/id/03C187CF-AD30-FF87-FF01-FF40FBEBFE45 |
|
treatment provided by |
Plazi |
|
scientific name |
Tatuidris tatusia Brown and Kempf 1968 |
| status |
|
Tatuidris tatusia Brown and Kempf 1968 View in CoL
Tatuidris tatusia Brown and Kempf 1968:187 View in CoL , their figures 1–4. Holotype and Paratype workers: El Salvador, La Libertad, 2 mi S. Quetzaltepec, VII-17-1961 (M.E. Irwin leg.) [holotype, UCDC] (image examined) [paratype, MCZC] (examined). Tatuidris kapasi Lacau and Groc , in Lacau et al. 2012:2, their figures 1–6. Holotype worker: Guyane Francaise, Montagne de Kaw, 0 4ο 38.21ˏ N; 0 52ο 17.36ˏ W, Alt. 260 m., ix.2008 (S. Groc, A. Dejean, and B. Corbara leg) [CPDC] (image examined). n. syn.
Worker, male and gyne diagnosis: With same characters as in the genus description ( Figures 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 ).
Pilosity variability. Currently, four striking pilosity patterns are known to occur within Tatuidris collections ( Figure 5 View FIGURE 5 ). Pilosity pattern A ( Figure 6 View FIGURE 6 ) consists of a mix of both long flexuous and short appressed setae. This is the most common pilosity pattern and the one that most resembles the type specimens from El Salvador and the gyne from Otongachi, Ecuador. Pilosity pattern B ( Figure 7 View FIGURE 7 ) is characterized by very short, fully appressed, and regular spaced setae arrayed homogeneously and equidistantly on the head, mesosoma, petiole, postpetiole and gaster. Pilosity pattern C ( Figure 8 View FIGURE 8 ) is characterized by dense lanose-looking setae. Pilosity pattern D ( Figure 9 View FIGURE 9 ) consists of short and uniform decumbent (strongly inclined but not fully appressed) setae scattered throughout the body.
DNA Barcodes. In total, 28 sequences (20 of them with full, 658bp, length) of the barcode gene were recovered from BOLD and included in the analyses ( Table 1 View TABLE 1 ). Among these sequences, 120 out of the 658 basepairs (18.23%) were variable. In general, specimens (excluding colony duplicates) presented very high levels of intraspecific pairwise differences (average = 5.36%, min = 0%, max=15.08%). The upper values of intraspecific pairwise differences encountered here are well above the 2% usually recovered for species differences in DNA barcode literature in general ( Ratnasingham and Hebert 2007), and above the percentages of average intraspecific variability reported previously for ants (e.g. between the 1.67% and 6.37% encountered for Anochetus and Odontomachus in Madagascar by Fisher and Smith 2008; see also Smith and Fisher 2009, Jansen et al. 2009). Pilosity patterns present among individuals (excluding pilosity pattern “C” for which no sequence was available) were not in similar clusters as depicted on the NJ tree ( Figure 10 View FIGURE 10 ). Instead, four different clusters (groups) with strong geographic structure were observed: G1, Mexico + Honduras + Guatemala; G2, Costa Rica + Nicaragua; G3, Ecuador (West of Andes); and G4, Ecuador (East of Andes/Amazon Basin) ( Figure 10 View FIGURE 10 ). Furthermore, across this geographic range, pairwise barcode divergence among specimens was significantly related to distance (R2=0.37, p<0.01, Figure 11 View FIGURE 11 ).
Size. Specimens of T. tatusia are small (average WbL = 0.62mm), but specimens can vary greatly in size, with larger specimens being twice as large as the smaller ones (min WbL = 0.45 mm, max WbL = 0.90 mm). Size variability within trap catches (possibly same colonies) may be considerable. For example, workers from one collection catch in Nicaragua (collection series MGB#1179) varied 30% in size (WbL from 0.65 to 0.85 mm). It is still unclear whether intra-colony size variation is due to the presence of morphological worker castes (e.g. minor and major castes) or continuous size variability. The PCA analysis revealed that most variability among specimens is related to size (proportion of variance explained by PC-1 = 0.915), with PC-2, PC-3 and PC-4 (e.g. shape) explaining little (0.033, 0.021 and 0.011, respectively) of total variation ( Table 2 View TABLE 2 ). Eye length loaded positively and contrasted against all other variables, in PC2. Tibia length and tibia width loaded negatively and contrasted against most other variables, in PC3. Size variability summarized in this analysis was not related to pilosity patterns ( Figure 12 View FIGURE 12 ). In general no PC correlated with pilosity patterns. Hand drawings courtesy of Natalie Clay ( Figure 12 View FIGURE 12 ).
Sculpture. The strength and depth of all sculpture patterns is accentuated in larger sizes. Collections from Nicaragua also tend to present more accentuated sculpture patterns. The head dorsum is usually smooth and shining, except for the area below eyes, which presents longitudinal carinae. The head vertex is covered with transverse carinulae. The lateral surface of the mandible is smooth and shining except for longitudinal superficial striae on the side that vary in depth. The antennal scape is shagreened and superficially areolate. The surface of the ventrolateral part of the pronotum varies strongly across specimens, from smooth and shining to strongly striate, or carinulate. The dorsum of the mesosoma has concentric carinulae and sometimes is slightly punctate. The mesopleuron is smooth and shining except for punctuations and areolae on the ventral margin. The propodeal declivity is smooth with fine transverse striae. The petiole and postpetiole are dorso-laterally strigulate. The gaster is mostly smooth and shiny but sometimes finely and sparsely strigulate.
Distribution. The genus Tatuidris is restricted to the Neotropics, but it has an ample distribution that spans from northern Mexico to central Brazil, French Guiana ( Lacau et al. 2012) and Amazonian Peru ( Figure 13 View FIGURE 13 ). No collections are known from the Caribbean, Galápagos, or other islands. Most specimens and collections are currently known to occur in localities west of the Andes, with more collections tending to occur towards Central America and Mexico. Most collections come from mountainside (pre-montane) areas at mid elevations (usually 800–1200m of altitude). Collections from lowland Amazonia are few.
Natural history. Little is known about the biology of the ant genus Tatuidris and, until recently, no observations of live specimens were registered. Details of a first collection event of a small live colony (3 workers and 4 gynes) by Dr. Thibaut Delsinne (pers. comm.) in a mid-elevation forest in southeastern Ecuador suggest that Tatuidris may well be a highly specialized predator, as colonies kept in captivity did not accept any food item offered to them. Food items rejected by the ants included live and dead termites, millipedes, mites, various insect parts, sugar water, tuna, biscuits, live and dead fruit flies ( Drosophila ), live springtails, live myriapods (Chilopoda and Diplopoda), live and dead Diplura, small live spiders, small live pseudoscorpions, one small snail, uncooked hen egg (i.e. piece of cotton wool soaked with fresh whisked hen egg; Brown 1977), ant larvae ( Gnamptogenys sp.), and live ant workers ( Cyphomyrmex sp., Brachymyrmex sp.). Potential food items (arthropods) for Tatuidris were taken from soil samples and Winkler samples (following Silva and Brandão 2010) collected at the site where Tatuidris was a priori determined to be present.
Further observations by T. Delsinne suggest that T. tatusia may be a sit-and-wait predator, as “both workers and gynes moved very slowly and were very clumsy. They often remained motionless during several tens of seconds or even several minutes when disturbed (either by my handling or by the contact with another arthropod). It is difficult to see them as powerful predators!” (pers. comm.). These observations were mainly performed at night, suggesting that T. tatusia may be nocturnal, a hypothesis also supported by collection patterns. For example, in the Río Toachi forest of Ecuador T. tatusia specimens tend to fall in pitfall traps, instead of Winkler sacs (Donoso and Ramón 2009). Because pitfall traps usually work 24-h, but Winkler sacs generally uses litter sifted during the day, then ants with nocturnal habits may be underrepresented in Winkler samples. The small eyes of Tatudris species provide further support for this hypothesis.
Other. The relative position of eyes is highly variable within the species. For example, eye location ranges from being completely within the antennal scrobes to completely outside the scrobes ( Figure 1 View FIGURE 1 b). In some cases (specimen J.Longino#2088-S) the eye itself is located outside the antennal scrobe, but the eye’s fossa is well marked and confluent with the antennal scrobe. In most specimens, the antennal carina bifurcates from the antennal scrobes and lies straight above the eyes. However, in specimens from Nicaragua (MGB#1179 colony collection), a strongly impressed antennal carina is present. In these specimens about 40% of the eye’s area lies within the antennal scrobes. In the gyne, only ~1/6 of the eye lies within the antennal scrobes. A depression sometimes forms in the integument in the sides of the propodeum, below the propodeal spiracle and above the metapleural gland. The depth of this depression varies among specimens and tends to be deepest in larger specimens.
Justification for the synonymy. A comprehensive analysis of the description of T. kapasi suggests that the morphological characters proposed by Lacau and Groc (2012) to separate T. kapasi from T. tatusia lie within the continuous variability encountered in Tatuidris specimens across the Neotropics. Two considerations were taken. First, the description of T. kapasi relies on a relatively large specimen where sculpture, body profile and body proportions are expected to vary the most. Diagnostic characters of T. kapasi (i.e. occipital border more concave, clypeus with the free margin nearly straight and laterally concave, dorsum of mesosoma mostly sculptured with concentric rugulae and carinulae, and dorsum of the petiolar and postpetiolar nodes with superficial concentric rugulae and carinulae) are all present to some degree in larger specimens among the collections reviewed in the present study ( Figures 6–9 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 ). Another diagnostic character of T. kapasi was denser pilosity without long setae (similar to Morph D), but setal patterns did not correlate with any morphology- or molecular-based grouping in this study. Second, the type locality of T. kapasi lies at a range limit of the genus, where morphological extremes might be expected.
Worker measurements (in mm) and indices: [average (min–max) of 10– 58 specimens]: AScL 0.46 (0.31, 0.67); AScW 0.24 (0.18, 0.36); CIx 129.03 (117.07, 137.93); EL 0.05 (0.03, 0.08); FL 0.43 (0.31, 0.70); FW 0.11 (0.08, 0.17); HL 0.59 (0.43, 0.88); HW 0.76 (0.56, 1.10); IAD 0.36 (0.25, 0.54); PL 0.16 (0.10, 0.24); PpL 0.16 (0.10, 0.25); PW 0.25 (0.18, 0.37); PpW 0.36 (0.26, 0.53); PPpIx 68.92 (58.14, 80.00); PrW 0.52 (0.38, 0.77); TiL 0.35 (0.27, 0.52); TiW 0.10 (0.06, 0.17); WbL 0.62 (0.45, 0.89); SL 0.42 (0.32, 0.51); SW 0.13 (0.11, 0.17); SI 329.82 (300.00, 360.00); REL 9.02 (5.41, 11.48).
Gyne measurements (in mm) and indices: (n=1) AScL 0.47; AScW 0.15; EL 0.20; FFS 0.09; FL 0.77; FW 0.20; HL 0.88; HW 1.28; IAD 0.56; Pl 0.22; PPL 0.28; PPW 0.72; PW 0.45; TL 0.69; TW 0.20; WingL 4.60; WbL 1.53; REL 22.63.
Male measurements (in mm) and indices: (n=1) EL 0.32; FFS 0.13; HL 0.66; HW 0.88; IAD 0.21; ScL 0.11; WingL 3.6; WbL 1.22; FL 0.9; REL 48.44.
Specimens examined: Elements in this list follow this order: COUNTRY, State or Province, Locality, elevation, coordinates, collector (collection code, habitat, method) [specimen code], [REPOSITORY].
BELIZE, 2.5 millas S Belmopan, S&J Peck (B-242, Limestone forest, ex: Berlese) [MCZC]. Caves Branch, S&J Peck (B-248, Hi Canopi Forest, ex: Berlese [MCZC]. BRAZIL, Amazonas, Manaus, Universidade do Amazonas, 03°05'36''S; 59°57'52''W, E Pereira Franken (Terra Firme: PLATO, ex: Pitfall) [INPA]. COLOMBIA, Magdalena, El Campano, 1300m, 11°07'N; 74°06'W, Phillip Ward (#7891-9, montane rainforest, ex: Sifted litter, leaf mold, rotten log) [QCAZ]. COSTA RICA, Rincon de la Vieja, Pailas, 1000m, 10.7887, –85.349, A Smith (10COSTA-0629 and 10COSTA-0630, Primary Forest) [AlexSmith]. Alajuela, Casa Eladio, Rio Peñas Blancas, 800m, 10°19' N; 84°43' W, J Longino (Wet forest, ex: Sifted leaf litter) [INBIOCRI002280066, INBIOCRI002280068 and INBIOCRI002280065] [JTLC]. Alajuela, El Aleman, Rio Peñas Blancas, 6,5 Km E Monteverde, 940m, 10°18' N; 84°45' W, J Longino (#0861-s, Wet forest, ex: Sifted leaf litter) [INBIOCRI002280059] [JTLC]. Heredia, 16km SSE La Virgen, 1100m, 10°16'N; 84°05'W [INB0003214087, INB0003214014, INB0003214104, INB0003214131, INB0003214232 and INB0003212497] [INBC]. Heredia, La Selva Biological Station-1, 50m, M Molina and H Garcia (AMI-1-W-006-05) [JTLC]. Heredia, La Selva Biological Station-2, 50m, 10.433, –84.0167, J Longino (All ants from Louisa Stark (OTS 89-1 student) study, ex: Litter and ground-nesting) [INBIOCRI001281194 and INBIOCRI001281181] [JTLC]. Heredia, La Selva Biological Station-2, 09°09' N; 079°51' W, Michael Kaspari (45196.01 and 45000.6) [MEK]. Puntarenas, Rio San Luis, 850m, 10°17' N; 84°48' W, J. Longino (Moist forest, ex: Sifted leaf litter on ground) [INBIOCRI002280062 and INBIOCRI002280063] [JTLC]. ECUADOR, Cotopaxi, 19 km ENE La Maná, 1100m, 00°53' S; 79°03' W, Phillip Ward (#11418-6, Second-Grown Rainforest, ex: Sifted leaf litter and logs) [QCAZ]. Pichincha, R.B. Maquipucuna, 1200m, 00°07'00''N; 78°38'06W, R Anderson (#99-208-6 and #99-208, Montane evergreen forest) [CASENT0423526 and CASENT0001968] [CASC]. Pichincha, Unión del Toachi-Otongachi, 850m, 00°21'05"S; 78°57'10"W, Donoso & Vieira (Bosque Secundario, ex: Pitfall) [18 workers and 1 gyne, MEKOU12079-84 and MEKOU12103-4] [QCAZ]. Zamora, Bombuscaro Numex, 950m, – 4.114972 S; – 78.96794 W, M Leponce (SPM_33796, Evergreen premontane rainforest, ex: Winkler sample) [QCAZ]. Zamora, Copalinga Private Reserve, 1000m, – 4.0912222 S; – 78.96069 W, T Delsinne and T. Arias-Penna (SPM_4130219, Secondary evergreen premontane rainforest, ex: Winkler sample) [QCAZ]. EL SALVADOR [Paratype and Holotype], 2 mi. S Quetzaltepec, ME Erwin [MCZ Paratype 31577] [MCZC]. GUATEMALA, Peten, Parq. Nac. Tikal, 270m, 17.232 N; – 89.623 W (Tropical Moist Forest) [CASC]. HONDURAS, Comayagua, PN Cerro Azul Meambar, 1120m, 14.887136; –87.89974 (LLAMA#Wa-C- 04-1-15, Ridgetop cloud forest, ex: Sifted leaf litter) [CASENT 0615399] [CASC]. MEXICO, Chiapas, 860m, 16.980 N; – 91.586 W, MG Branstetter (#856, Mesophil forest) [QCAZ]. Chiapas, 12 mi NW Ocozocoautla, 400m, A Newton (ex: Berlese) [MCZC]. Chiapas, 6kmSW Ocosingo, 1400m, 16.867221 N; – 92.0787132 W, R Anderson (# 91–116, Forest litter, ex: Berlese) [CASENT 0603397] [INBC]. Chiapas, Lago Metzabok, 575m, 17.124, –91.636, MG Branstetter (Lowland wet forest) [CASC]. Oaxaca, Mirador Grande, 990m, 17.89844, –96.36253, MG Branstetter (#1405) [QCAZ]. Tamaulipas, El Cielo, 870m, 23.276, –99.276, MG Branstetter (#1465) [QCAZ]. Ve r a c r u z, Los Tuxtlas, Ejido-López Mateos, 50m, 18°24'56''N; 94°56'53''W, P Rojas (LM7S-H1, LM42S-H4, LM42S-H5 and LM21A-H3, Selva alta perennifolia, ex: Winkler) [IEXM]. Veracruz, Los Tuxtlas, Ejido-López Mateos, Ejido 2, 50m, 18°24'56''N; 94°56'53''W, P Rojas (LM42S- H3, Selva alta perennifolia, ex: Winkler) [IEXM]. Veracruz, Los Tuxtlas, Volcán S. M. Pajapan, 510m, 18°16'00''N; 94°46'71''W, A Cartas (847d, Selva mediana subperennifolia, ex: Berlese) [IEXM]. NICARAGUA, Matagalpa, RN El Musún, 4.8km NNW Rio Blanco, 1170m, 12°58.4'N; 085°14'W, M.G.Branstetter (#1179, Mesic forest, ex: Sifted leaf litter) [QCAZ]. PANAMA, Chiriqui, 20,4 Km North San Felix, R. Anderson (#17768_1, Wet mountain forest, ex: Litter sample) [WEMC]. Chiriqui, Alto Lino, 3800m, HG Real [Male, CASENT 0102681] [CASC]. Chiriqui, La Fortuna, Finca La Suisse, 1200m, R. Anderson (#17838, #17787, #17788, #17789, #17790, #17838 and #17839 [WEMC]. PERU, Cuzco, La convención Province, 4 km S Camisea River. Campamento Cashiriari-2, Plot 1, 579m, 11°51'51.3''S; 72°46'45.6''W, J Santisteban et al. (#38, Primary Rainforest, hilly terrain, ex: Winkler Trap) [MUSM-ENT 0201599/ANTWEB-CASENT 0178882] [MUSM].




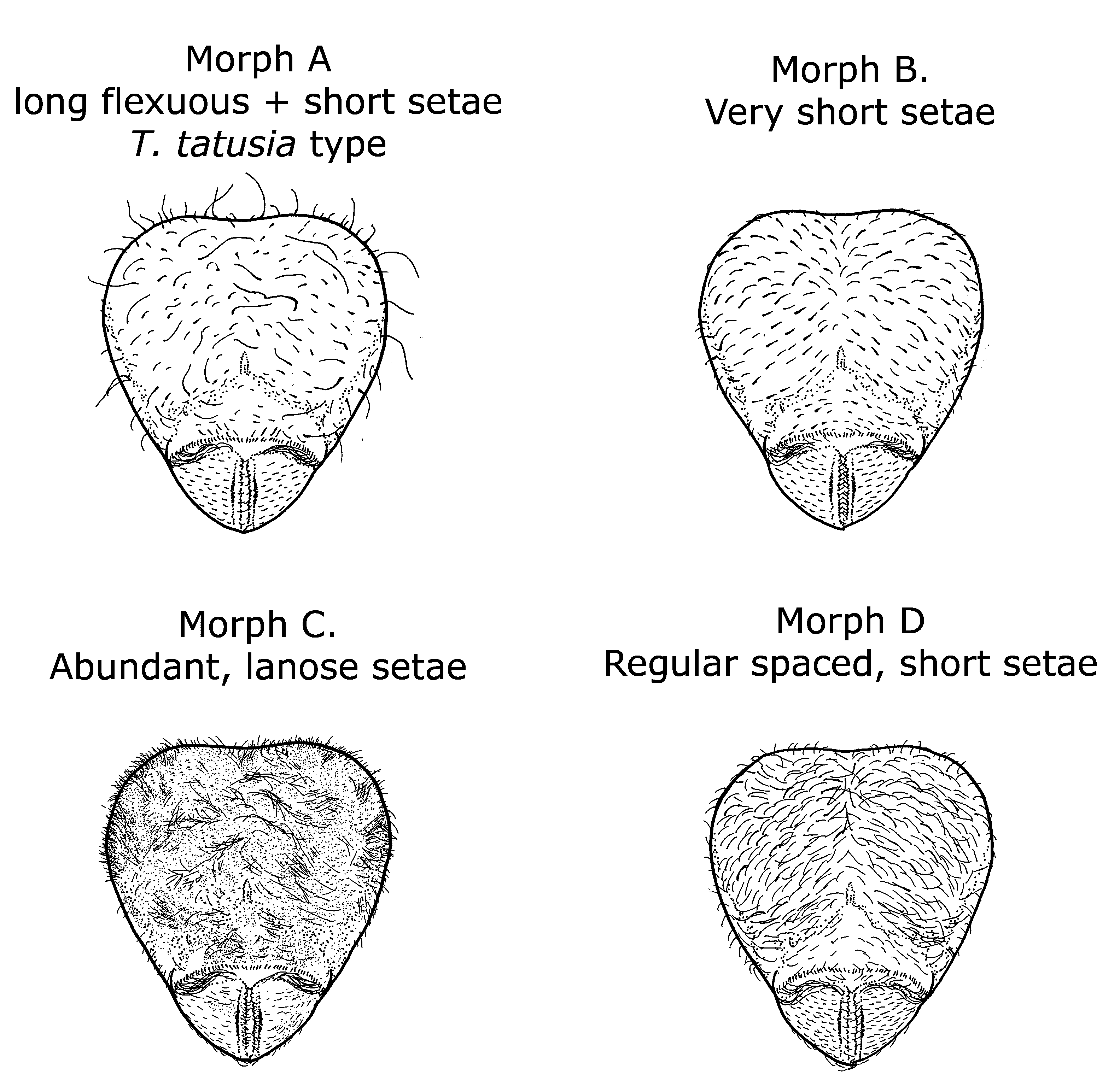





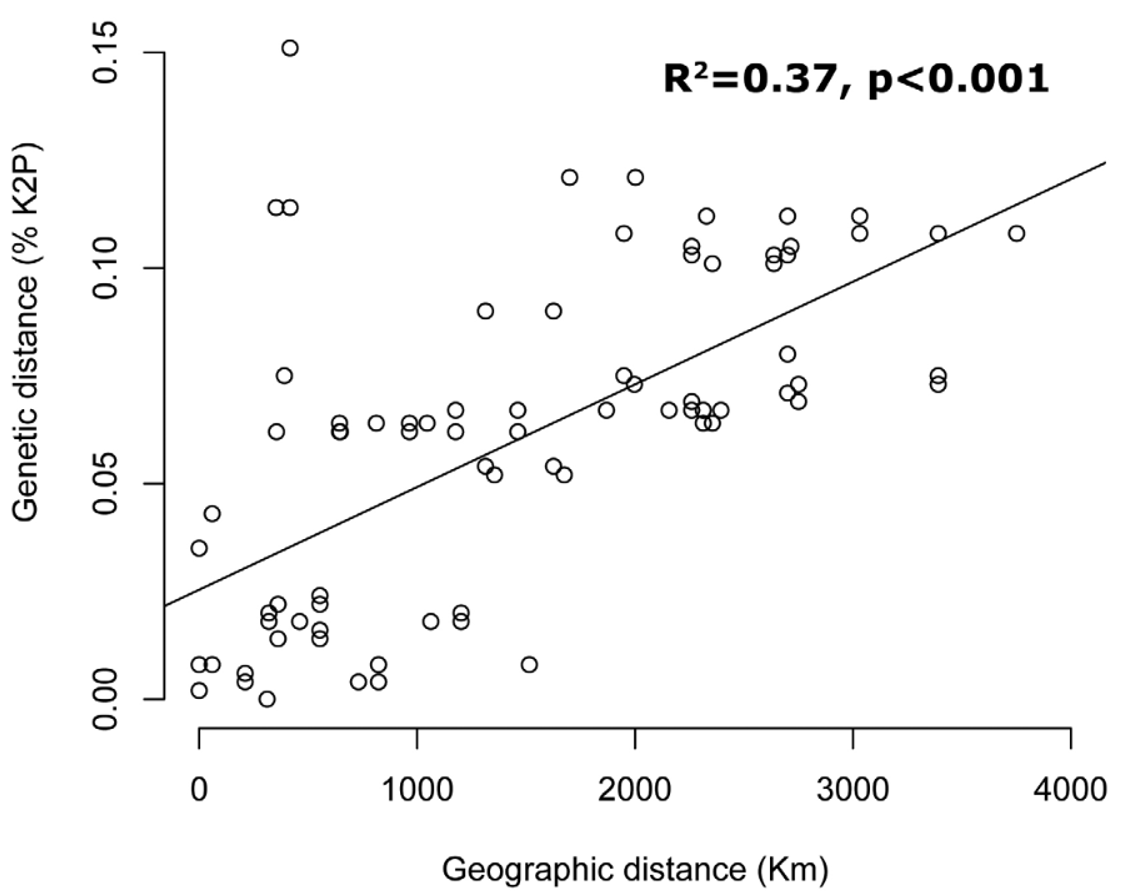
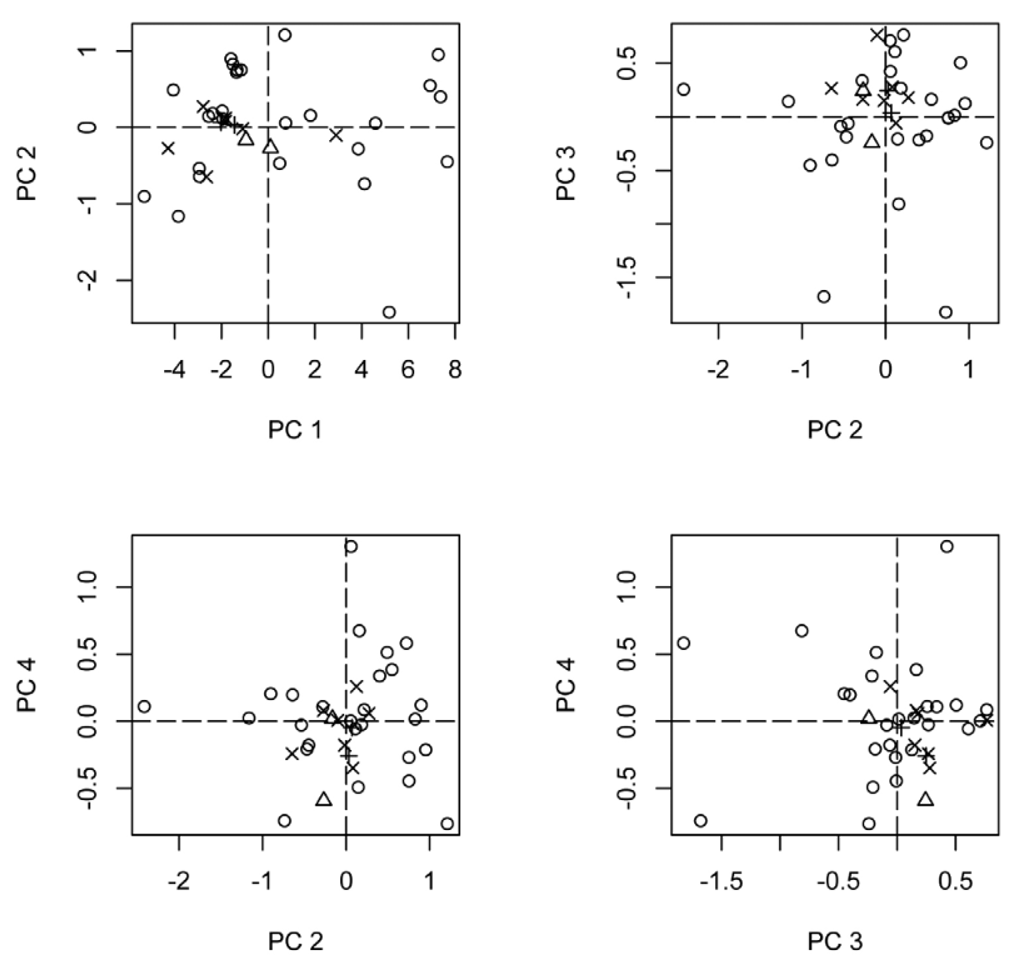
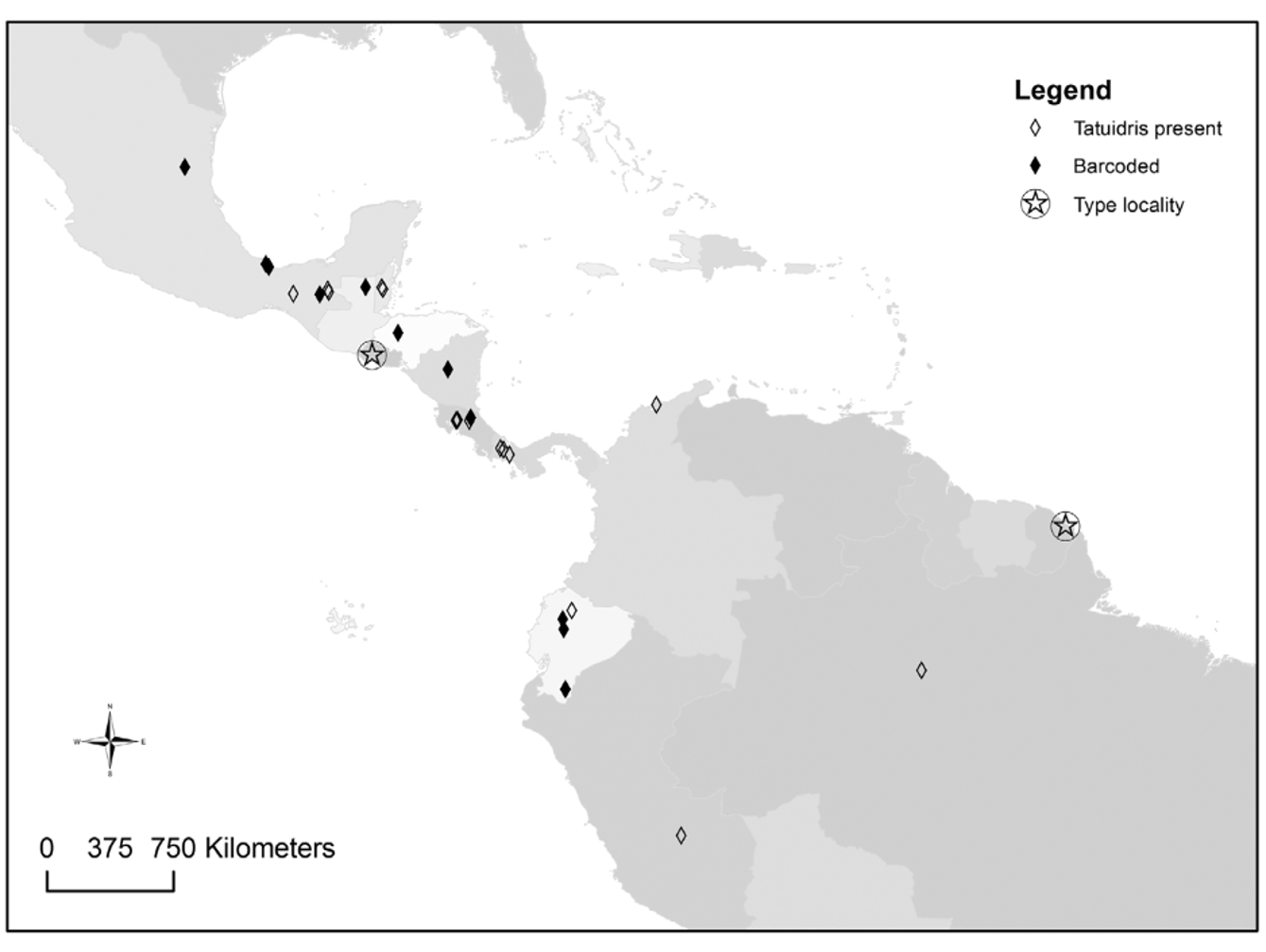
TABLE 2. First four PCA coefficients for morphological variables retained in this study. PC eigenvalues (% of variance explained) are also given.
| Trait | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| AScL | 0.2737 | –0.0619 | 0.0329 | 0.2525 |
| AScW | 0.2730 | –0.1618 | –0.0559 | 0.1040 |
| EL | 0.2135 | 0.9362 | –0.1003 | –0.0722 |
| HL | 0.2761 | –0.0490 | 0.0880 | 0.2140 |
| HW | 0.2780 | –0.0043 | 0.0557 | –0.0271 |
| IAD | 0.2749 | –0.1594 | 0.0246 | 0.1754 |
| PL | 0.2679 | –0.1677 | 0.1582 | –0.4504 |
| PpL | 0.2643 | 0.0462 | 0.4339 | –0.3384 |
| PW | 0.2739 | –0.0537 | 0.1803 | –0.1468 |
| PpW | 0.2758 | 0.0473 | 0.0436 | 0.1971 |
| PrW | 0.2767 | 0.0585 | –0.0261 | 0.0450 |
| TiL | 0.2440 | –0.1402 | –0.8193 | –0.4051 |
| TiW | 0.2672 | –0.0193 | –0.2126 | 0.5268 |
| WbL | 0.2752 | –0.0823 | 0.0967 | –0.1464 |
| Eigenvalue | 0.9154 | 0.0334 | 0.0211 | 0.0112 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Tatuidris tatusia Brown and Kempf 1968
| Donoso, David A. 2012 |
Tatuidris tatusia
| Lacau 2012: 2 |
| Brown 1968: 187 |